 Reproduced, with permission, from:
Dobson, A., and R. Carper. 1992. Global warming and potential changes in host-parasite and disease-vector relationships. In Global warming and biodiversity, ed. R. L. Peters and T. E. Lovejoy. New Haven, CT: Yale University Press.
Reproduced, with permission, from:
Dobson, A., and R. Carper. 1992. Global warming and potential changes in host-parasite and disease-vector relationships. In Global warming and biodiversity, ed. R. L. Peters and T. E. Lovejoy. New Haven, CT: Yale University Press.
ANDREW DOBSON AND ROBIN CARPER
Parasitology has always been a discipline in which purely academic studies of the evolution of parasites and their life cycles have progressed as a necessary complement to the study of the pathology and control of the major tropical diseases of humans and their livestock. Indeed, the most striking feature of parasitology is the diversity of parasites in the warm tropical regions of the world and the frightening levels of debilitation and misery they cause. Determining how long-term climatic changes will affect the distributions of different parasites and pathogens at first seems a daunting task that almost defies quantification. Nevertheless. as parasitologists have always been concerned with the influence of climatological effects on different parasite species, it is possible to begin to speculate on the ways that global warming might affect the distributions of some specific tropical diseases. Similarly, the study of parasite population dynamics has developed within a solid theoretical framework (Anderson and May 1979. May and Anderson 1979). This permits the development of quantitative speculation in more general studies concerned with how parasite-host interactions may respond to perturbation.
This chapter addressed both general questions about the response of parasite-host systems to long-term climatic changes and the specific response of one particular pathogen, Trypanosoma, to the changes in climate predicted for the next hundred years.
Current estimates suggest that parasitism of one form or another may be the most common life-history strategy in at least three of the five major phylogenetic kingdoms (May 1988, Toft 1986). The enormous array of pathogens that infect humans and other animals may be conveniently divided on epidemiological grounds into microparasites and macroparasites (Anderson and May 1979, May and Anderson 1979). The former include the viruses, bacteria, and fungi and are characterized by their ability to reproduce directly within individual hosts, their small size and relatively short duration of infection, and the production of an immune response in infected and recovered individuals. Mathematical models examining the dynamics of microparasites divide the host population into susceptible, infected, and recovered classes. In contrast, the macroparasites (the parasitic helminths and arthropods) do not multiply directly within an infected individual but instead produce infective stages that usually pass out of the host before transmission to another host. Macroparasites tend to produce a limited immune response in infected hosts; they are relatively long-lived and usually visible to the naked eye. Mathematical models of the population dynamics of macroparasites have to consider the statistical distribution of parasites within the host population.
A second division of parasite life histories distinguishes between those species with monoxenic life cycles and those with heteroxenic life cycles. The former produce infective stages that can directly infect another susceptible definitive host individual. Heteroxenic species utilize a number of intermediate hosts or vectors in their transmission between definitive hosts. The evolution of complex heteroxenic life cycles permits parasite species to colonize hosts from a wide range of ephemeral and permanent environments, while also permitting them to exploit host populations at lower population densities than would be possible with simple direct transmission (Anderson 1988, Dobson 1988, Mackiewicz 1988, Shoop 1988). However, heteroxenic life cycles essentially confine the parasite to areas where the distributions of all the hosts in the life cycle overlap.
Shifts in the distribution of these host species due to climatic changes, will therefore be important in determining the areas where parasites may persist and areas where parasites may be able to colonize new hosts.
Climatic changes are likely to have different effects on aquatic and terrestrial environments (chapter 24). The heteroxenic life cycles of some parasite species often allow them to utilize hosts sequentially from either type of habitat. It is thus important to determine the different responses of the terrestrial and aquatic stages of a parasite's life cycle to climatic change. That, along with an examination of other parasite responses to climatic change, demands a quantitative framework within which to discuss parasite life-history strategies.
The complexities of parasite host population dynamics may be reduced by the derivation of expressions that describe the most important epidemiological features of a parasites life cycle (Anderson and May 1979, May and Anderson 1979, Dobson 1988). Three parameters are important in describing the dynamics of a pathogen: the rate it will spread in a population, the threshold number of hosts required for the parasite to establish, and the mean levels of infection for the parasite in the host population.
Basic reproductive rate of a parasite, Ro: The basic reproductive rate, Ro, of a microparasite may be formally defined as the number of new infections that a solitary infected individual is able to produce in a population of susceptible hosts (Anderson and May 1979). In contrast, Ro for a macroparasite is defined as the number of daughters that are established in a host population following the introduction of a solitary fertilized female worm. In both cases the resultant expression for Ro usually consists of a term for the rates of parasite transmission divided by an expression for the rate of mortality of the parasite in each stage in the life cycle (Dobson 1989). Increases in host population size or rates of transmission tend to increase Ro, and increases in parasite virulence or other sources of parasite mortality tend to reduce the spread of the pathogen through the population.
Threshold for establishment, HT: The threshold for establishment of a parasite, HT is the minimum number of hosts required to sustain an infection of the pathogen. An expression for HT may be obtained by rearranging the expression for Ro to find the population density at which Ro equals unity. This may be done for both micro- and macroparasites with either simple or complex life cycles. The resultant expressions suggest that changes in the parameters that tend to increase Ro tend to reduce HT and vice versa. Although many virulent species require large populations to sustain themselves, reductions in the mortality rate of transmission stages may allow parasites to compensate for increased virulence and maintain infections in populations previously too small to sustain them.
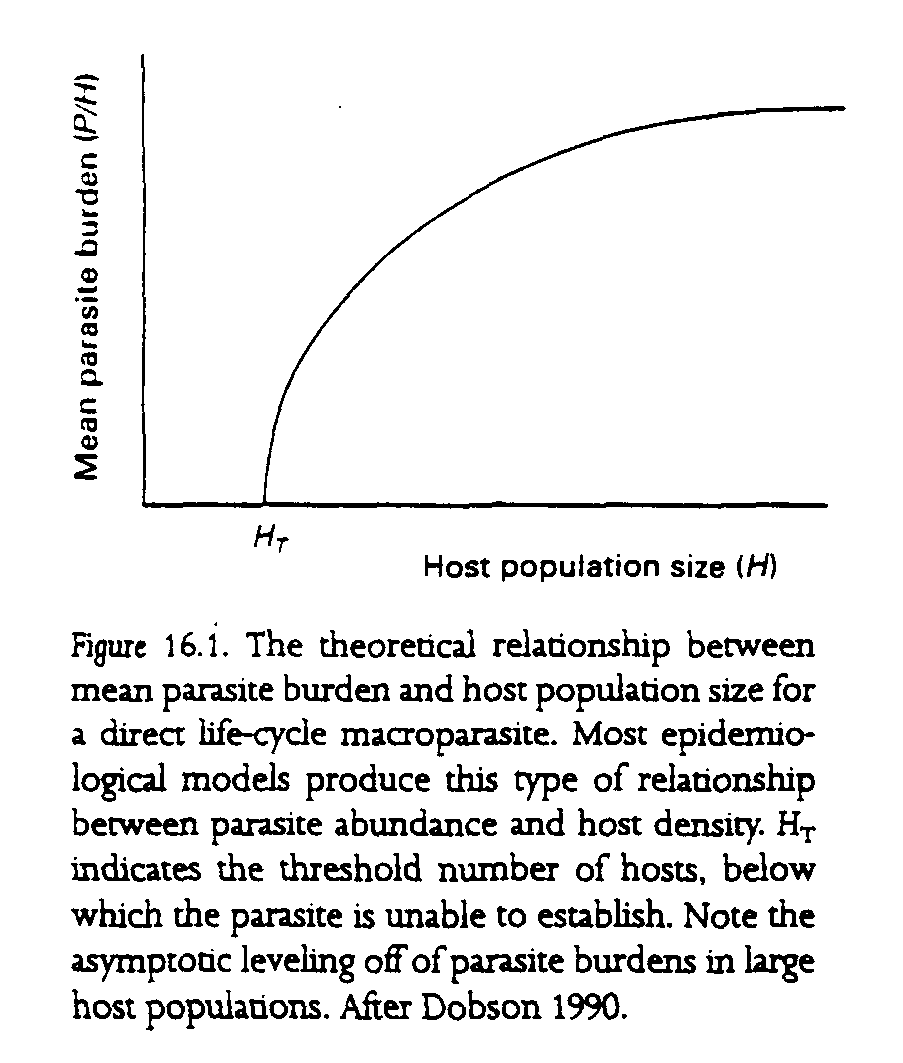
Mean prevalence and burden at equilibrium: It is also possible to derive expressions for the levels of prevalence (proportion of the hosts infected) and incidence (mean parasite burden) of parasites in the host populations. In general, parameters that tend to increase Ro also tend to give increases in the proportion of hosts infected by a microparasite and increases in the mean levels of abundance of any particular macroparasite (Anderson and May 1979, May and Anderson 1979, Dobson 1988). Most important, increases in the size of the host population usually lead to increases in the prevalence and incidence of the parasite population (fig. 16.1).
These expressions, which characterize the most important features of a parasite's interaction with its host at the population level, can be used to ascertain how parasites with different life cycles will respond to long-term climatic changes. This may best be undertaken by determining which stages of the life cycles are most susceptible to climatic variation and by quantifying the response of those stages to climatic change.
The physiology of adult parasites is intimately linked with the physiology of their hosts. Providing the hosts can withstand environmental changes, it seems unlikely that the within-host component of the parasite life cycle will be significantly affected. However, any form of increased stress on the host may lead to increases in rates of parasite-induced host mortality (Esch et al. 1975). In the absence of data from the specific experimental studies that could throw considerable light on these relationships, this study will concentrate on the effect of changes in meteorological factors on the free-living infective stages of different groups of parasites.
Several detailed laboratory studies have examined the effect of temperature on the transmission success of parasites with aquatic infective stages. The parasitic trematodes are probably the most important class of parasites to utilize an aquatic stage for at least part of their life cycle. The data presented in figure 16.2 are for an echinostome species that is a parasite of ducks. Increased temperature leads to increased mortality of the larval infective stages of the parasite, also leads to increased infectivity of the larval stage. The interaction between larval infectivity and survival means that net transmission efficiency peaks at some intermediate temperature but remains relatively efficient over a broad range of values (16deg.-36deg.C for Echinostoma liei cercariae; fig. 16.2). These synergistic interactions between the different physiological processes determining survival and infectivity allow the aquatic parasites to infect hosts at a relatively constant rate over the entire spectrum of water temperatures that they are likely to experience in their natural habitats (Evans 1985).
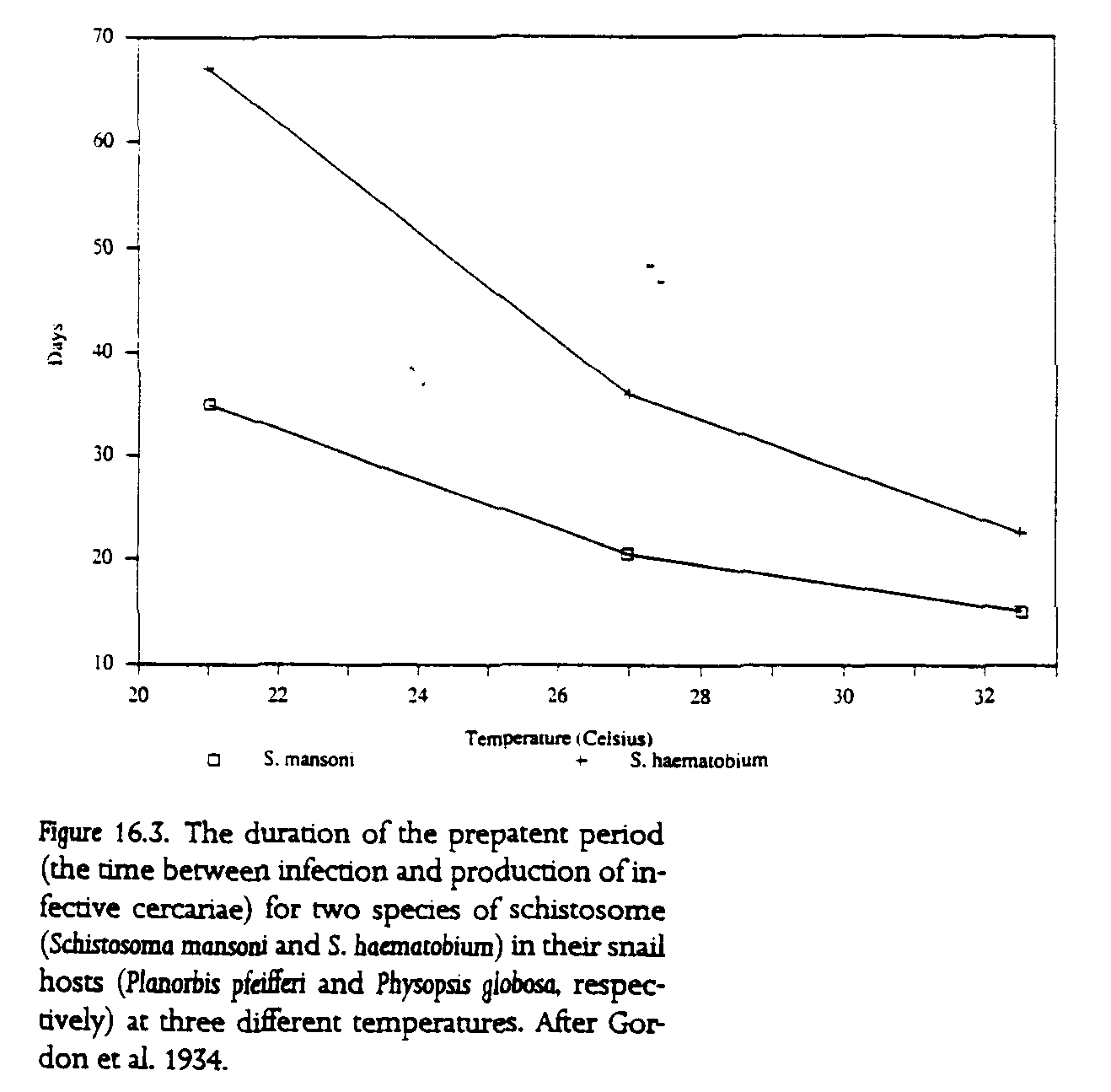
The effect of temperature on the developmental rate of parasites in both aquatic and terrestrial hosts has been examined for several of the major parasites of humans in the tropics. In contrast to the effect on transmission efficiency, increases in temperature usually lead to reduced development times for parasites that utilize poikilothermic hosts (fig. 16.3). As with many physiological processes, a 10deg. increase in temperature seems to lead to a halving of the developmental time. This may allow parasite populations to build up rapidly following increases in temperature.
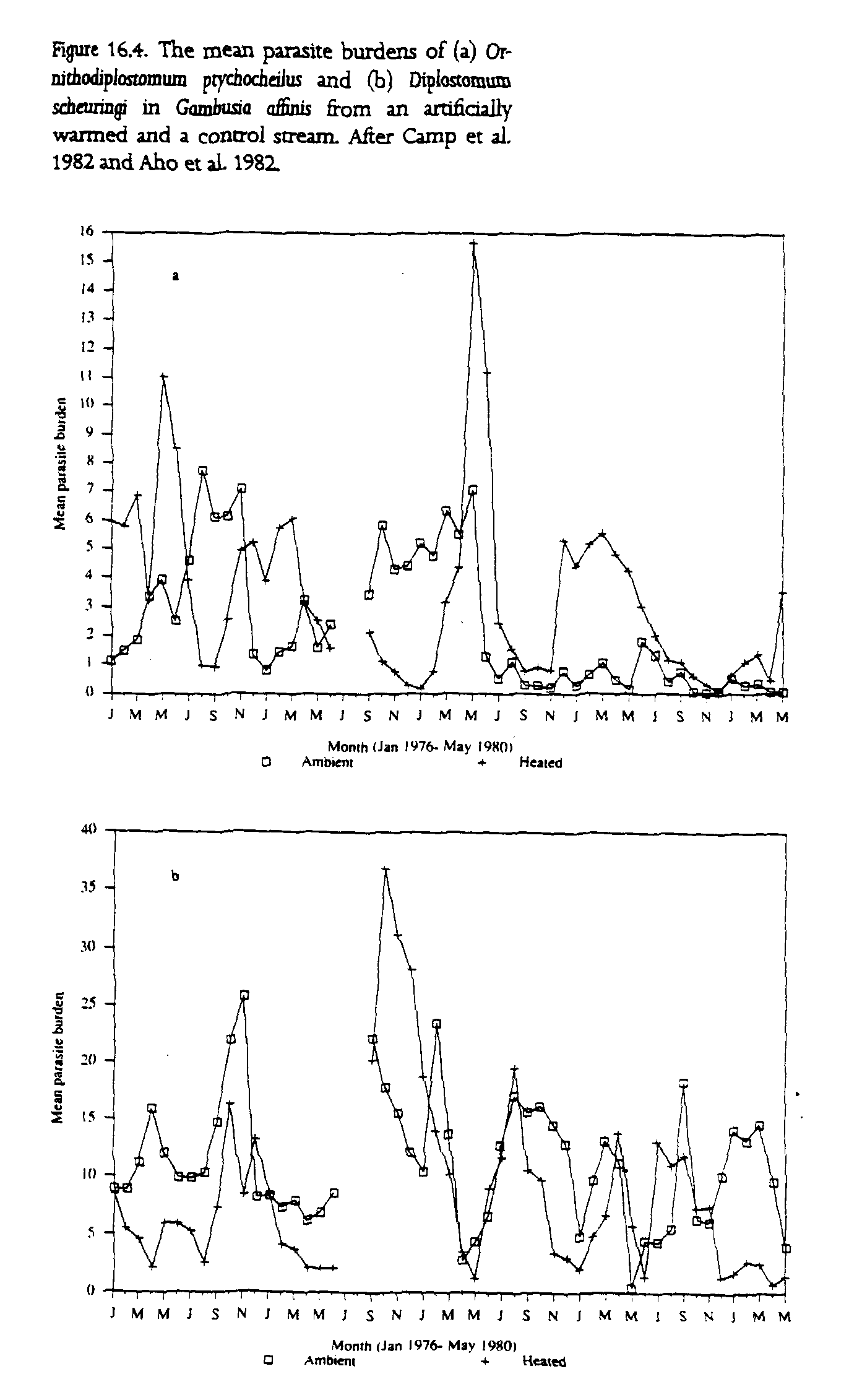
The expressions for Ro and HT, derived in the first part of this chapter, suggest that increases in transmission efficiency and reductions in development time induced by temperature changes allow parasites to establish in smaller populations and grow at more rapid rates. This is observed to some extent in a pair of long-term studies that compare the parasite burdens of mosquito fish (Gambusia affinis) populations in artificially heated and control sections of the Savannah River in South Carolina The data for the trematode Ornithodiplostomum ptychocheilus show significant differences between heated and ambient sites during the earlier period of the study when temperature differences were most pronounced. Infection by the parasites starts several months earlier each year in the thermally altered sites (fig. 16.4). However, infection rates decline in the summer in the artificially heated sites when populations of hosts decline in response to high water temperatures (Camp et al. 1982). This effect may be compounded by the movement of the waterfowl that act as definitive hosts for the parasite. These birds tend to prefer the warmer water in winter and cooler water in the summer. Similar but less clearly defined patterns are observed in the data for Diplostomum scheuringi from the same site (Aho et al. 1982).
These studies illustrate the important role of host population density in the response of a parasite's transmission rate to thermal stress, while also demonstrating the ability of parasites to capitalize on improved opportunities for transmission and to establish whenever opportunities arise. Obviously the data are open to several interpretations, but they do emphasize the importance of long-term experiments in determining the possible effects of global warming on the distribution of parasites.
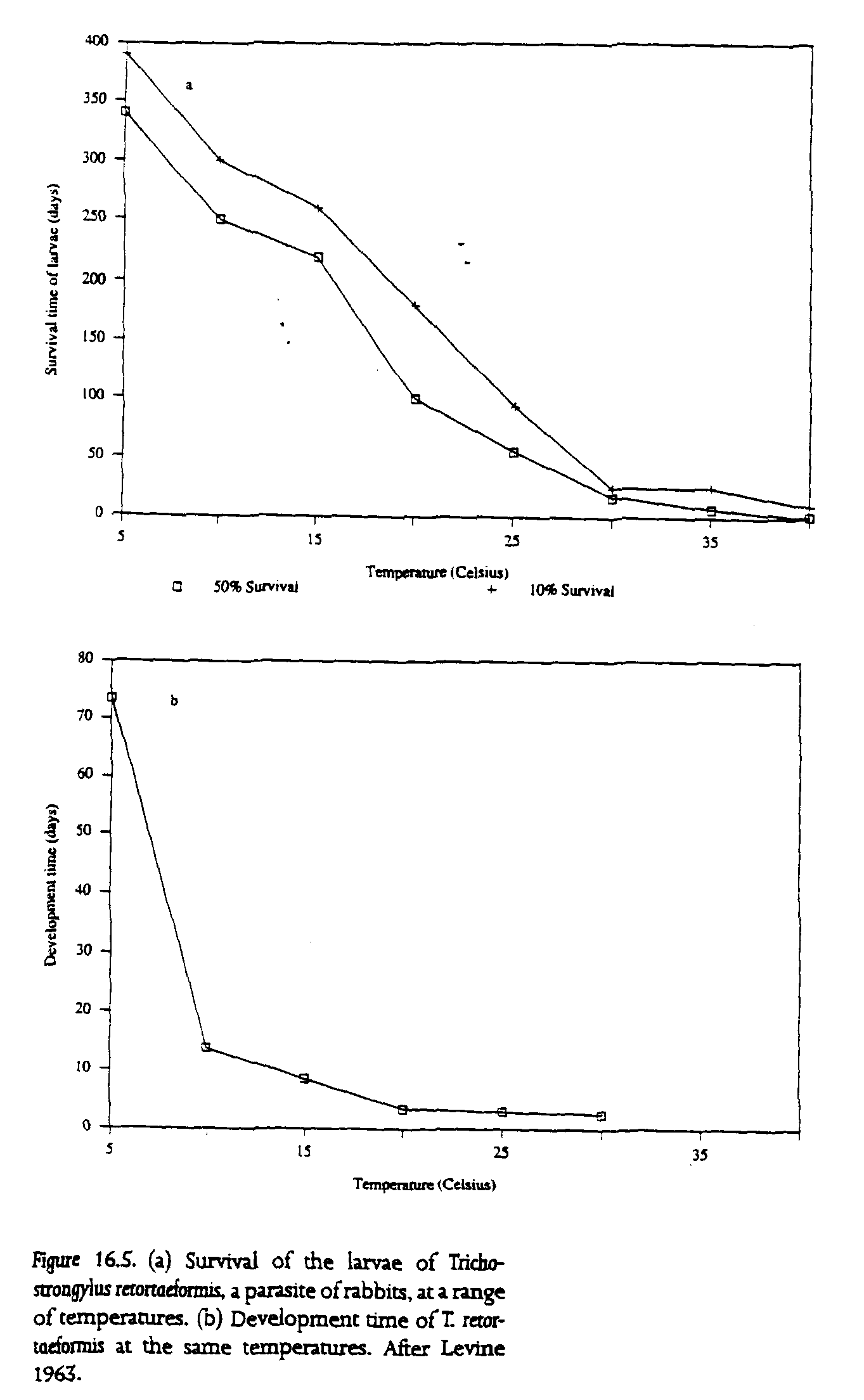
The survival rates of the infective stages of the parasites of most terrestrial species tend to decrease with increasing temperature (fig. 16.5a). Although little evidence is available to determine how the infectivity of these larvae is affected by temperature, rates of larval development tend to increase with increasing temperature (fig. 16.5b). These two processes again interact synergistically---as an increase in temperature depresses survival, development speeds up---allowing the parasite to establish at a broad range of environmental temperatures. In contrast to parasites that utilize aquatic hosts, parasites of terrestrial hosts have transmission stages that are susceptible to reduced humidity, and these stages are highly susceptible to desiccation (Wallace 1961). To compensate for reduced opportunities for transmission during periods of severely adverse climate, parasites of terrestrial hosts have evolved adaptations such as hypobiosis, the ability to remain in a state of arrested development within the relatively protected environment provided by their hosts until such time as transmission through the external environment proves more effective. Terrestrial nematodes, for example, can arrest their development. This ability is a heritable trait and one that seems to adapt rapidly to different climatological and management regimes (Armour and Duncan 1987).
Because interactions between temperature and humidity seem to be of major importance in constraining the geographical range of many of the pathogens that infect domestic livestock, a considerable body of data exists concerning the relation between meteorological conditions and parasite outbreaks (Gordon 1948, Kates 1965, Levine 1963. Ollerenshaw 1974, Wilson et al. 1982). Indeed the parasitologists of the 1950s and 1960s firmly believed that climate determined the distribution of a parasite species, while weather influenced the timing of disease outbreaks. Large-scale research programs were designed to forecast disease outbreaks in different areas and to recommend the best time to administer control measures.
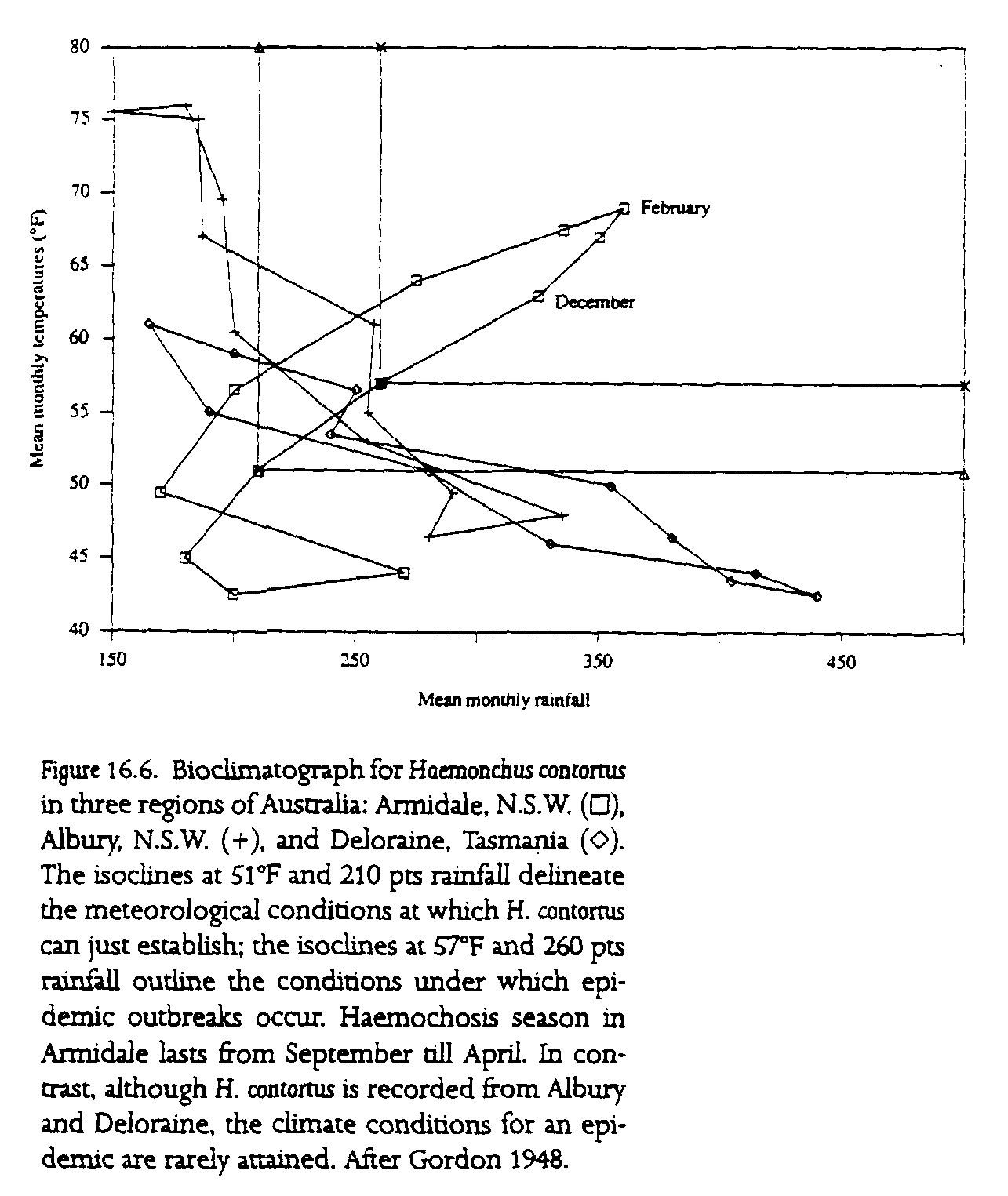
One way of depicting the interaction between disease outbreaks and climate was through bioclimatographs (fig 16.6). The use of these diagrams for monitoring parasite outbreaks was originally suggested by H. M. Gordon (1948) in a study of the sheep nematode Haemonchus contortus (the barber's pole worm). Bioclimatographs are constructed by plotting the climatological conditions under which a parasite is able to exist and under which outbreaks occur onto a graph of mean monthly temperature and rainfall. When that plot is compared with the observed mean weather data for a specific geographical location, it is possible to determine the time of year when outbreaks of the parasite are likely. Although the initial production of a diagram requires a long-term study of the parasite in any region, once the conditions for establishment and optimal development have been described, then extrapolations may be made to other regions for which only the climate data are available. Thus Gordon (1948) was able to use his data for H. contortus in Armidale, New South Wales, where outbreaks occur from October to May, to explain why outbreaks rarely occurred in other regions such as Albury, N S.W., and Deloraine, Tasmania.
N. D. Levine (1963) reviewed and extended the use of bioclimatographs to define and explain the distribution and seasonal incidence of a variety of gastrointestinal parasites of sheep and cattle. Being based on mean temperature and rainfall data, bioclimatographs are usually only partially successful in predicting parasite outbreaks in any specific year. Similarly, bioclimatographs are seldom derived from laboratory determinations of a parasite's development constraints, because the climate conditions experienced by the parasite larvae in the soil are often different from those measured by the local weather station. However, bioclimatographs remain useful tools for determining whether a parasite will establish in a region. They may prove invaluable in determining whether long-term climatic changes will permit specific parasites of domestic livestock to establish in regions where they are not at present a problem.
Our focus so far on parasitic helminths reflects the available literature. Data on the effects of temperature, humidity, and ultraviolet light on the survival and infectivity of viral and bacterial transmission stages have been hard to locate, possibly because work with this material is beset with technical difficulties. There are, however, data suggesting that the development time of microparasite infections depends on ambient temperature, and there is evidence that the infectivity of some vector-transmitted pathogens is determined by the temperature at which their insect hosts are raised (Ford 1971:104). Temperature may also indirectly affect transmission rates by altering the behavior of insect vectors.
Trypanosomiasis, or sleeping sickness, is one of the major diseases of humans and their domestic animals in Africa (Ford 1971). The disease is of particular importance to conservation in Africa as its presence may exclude humans and their domestic livestock from areas where wild animals act as a reservoir of the disease (Molyneux 1982, Rogers and Randolph 1988). The pathogen may be classified as a microparasite; it is transmitted by an insect vector, the tsetse fly (Glossina spp.). D. J. Rogers and S. E Randolph have made an extensive study of the meteorological conditions that determine the distribution of three species of tsetse flies, Glossina morsitans, G. palpalis, and G. tacinoides (Rogers 1979, Rogers and Randolph 1986). Their study is complemented by two models of the dynamics of the different Trypanosoma species, one by Rogers (1988) and one by P.J.M. Milligan and R D. Baker (1988). The former derives expressions for Ro and HT that provide some useful general insights into the processes that are most important in determining the conditions that allow the pathogen to establish; the latter develops a more specific analytical model for trypanosomiasis based on detailed parameter estimates from a study of Trypanosoma vivax in Tanzania.
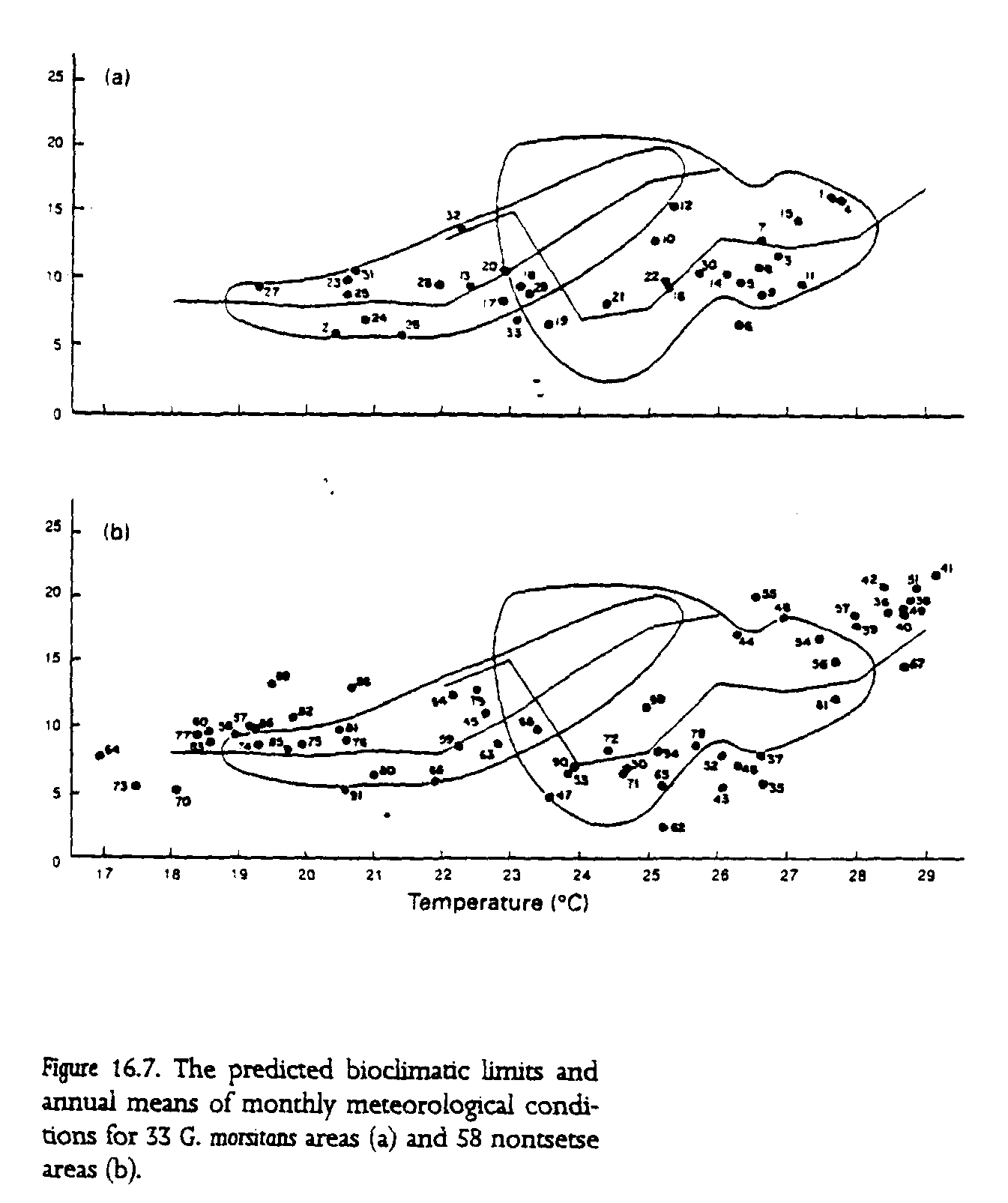
Rogers's (1979) analysis of the bioclimatic tolerances of tsetse flies may be used to determine how predicted patterns of climate change in tropical Africa might affect the distribution of tsetse flies and trypanosomiasis. Using data from several long-term studies of two subspecies of tsetse flies in Nigeria (Glossina morsitans submorstans) and Zambia (G. m. morsitans), Rogers shows that the mean monthly density-independent mortality rates for these flies are most closely related to mean monthly saturation deficit (an index of humidity) and, to a lesser extent, mean monthly temperature. Those analyses allow Rogers to identify an environmental optimum for each subspecies of G. morsitans. When the data for 91 sites throughout tropical Africa are examined in terms of these climatological conditions, 94% of the sites within the present known distribution of G. morsitans fall within the predicted bioclimatic limits, while only 50% of nontsetse areas do so (fig. 16.7). These data can be used to compare the present distribution of G. morsitans with the possible distribution given a mean 2deg. increase in temperature for sub-Saharan Africa (fig. 16.8). Because the bioclimatic data correlate better with the presence of G. morsitans than they do with its absence, greater confidence may be placed in the prediction for where G. morsitans may decrease in abundance than for regions where it might establish. Keeping that in mind, the analysis suggests that G. morsitans may become less common in West Africa and across the main sub-Saharan zone of central Africa This pattern may be matched by a spread farther south of the vector and its parasite in East Africa
Although the approach we have adopted in this analysis is rather coarse, the data are available to make more sophisticated and detailed analyses for this and other pathogens. From a conservation perspective it remains important to determine to what extent trypanosomiasis is at present maintaining areas as refuges for wild animals by excluding humans and their livestock (Molyneux 1982, Rogers and Randolph 1988). If a change of climate reduces tsetse levels, then pressure for the exploitation of the areas would increase with their subsequent loss as a wildlife refuge.
So far we have concentrated on simple one-host, one-parasite relationships, a sensible approach to systems that are dominated by one particularly prevalent pathogen. Many host populations. however, maintain a community of several parasite species. The diversity of such a community and the abundance of its constituent parasite species are intimately linked not only to the density of the host population but also to the presence of other host species that act as reservoirs for other parasite species.
It is possible to extend the basic one-host, one-parasite models to examine the dynamics of more complex communities (fig. 16.9). Preliminary analysis of models for such communities suggests that parasite species diversity is a direct function of host density and that the relative abundance of each parasite species is determined more by the parasite's life-history attributes that determine its transmission success than by interactions with other parasite species (Dobson 1986, 1989). This suggests that changes in host density due to changes in meteorological conditions will be crucial in determining the diversity of the community of parasites supported by the hosts. Increases in the density of some hosts will allow them to support a more diverse parasite fauna, while decreases in the density of other hosts will reduce the diversity of their parasite community.
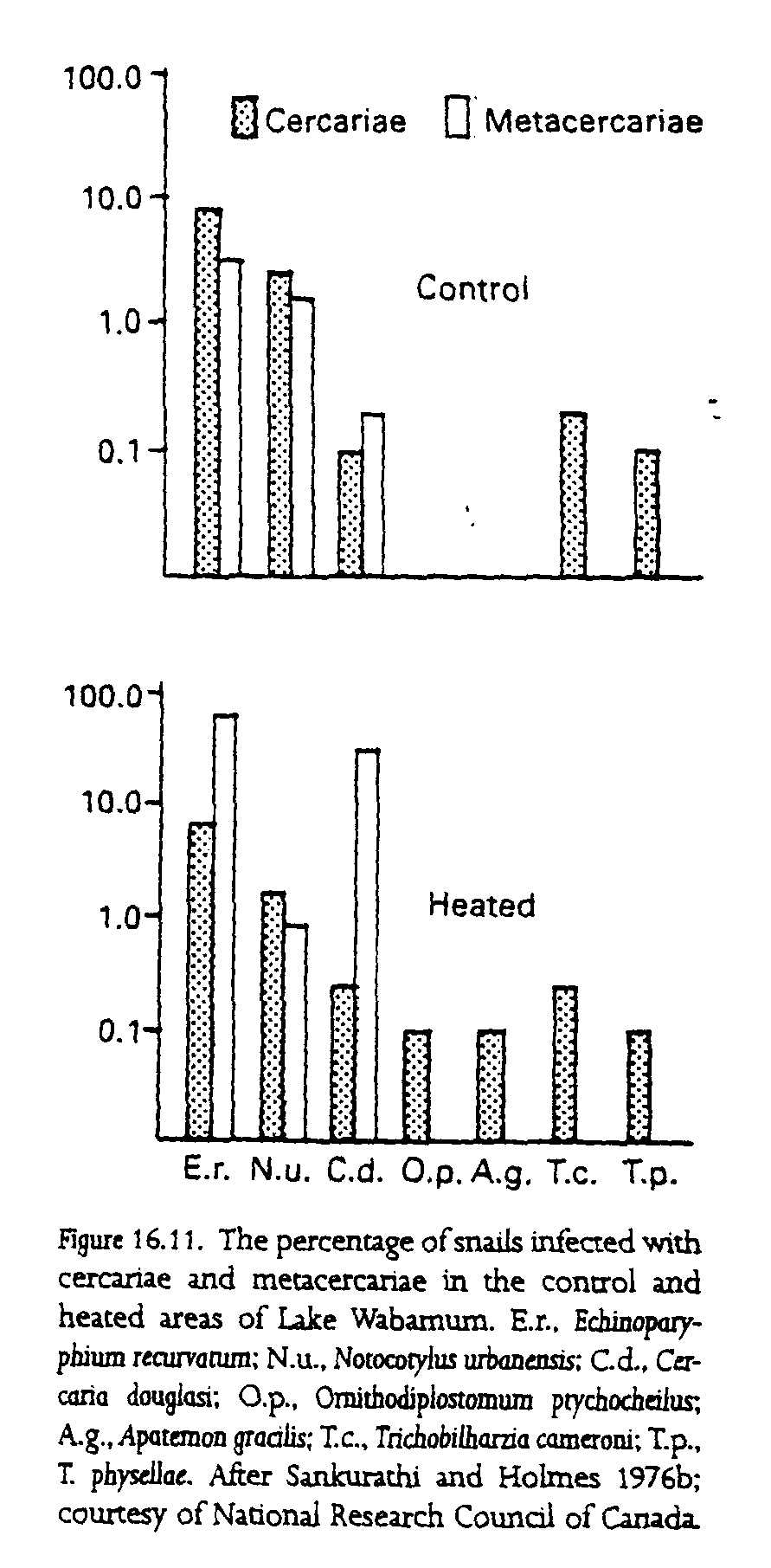
A study comparing the effects of artificial heating on the parasite fauna of an aquatic snail presents some corroborative evidence in support of this model. C. S. Sankurathi and J. C. Holmes (1976a,b) studied a population of Physa gyrina and its parasites and commensals in Lake Wabamun in Alberta, Canada. A section of the lake was used for cooling by a power station and consequently was warmer than the rest of the lake and relatively free of ice in winter. The effects on the population of snails were pronounced when both density and population structure are compared for heated and control sites, with population density often several orders of magnitude higher in the heated areas (fig. 16.10). That, and the continual presence of the vertebrate definitive hosts in the parasites life cycle, allowed a considerable increase in both the prevalence and diversity of the parasite community living in the snail population (fig. 16.11). The increased water temperature also had a detrimental effect on the two species of commensal chaetogasters that live in the mantle of the snails. Laboratory experiments showed that these commensals live as predators, attacking and ingesting the infective stages of parasites that try to infect their snail host (Sankurathi and Holmes 1976b). When the temperature rises, the chaetogasters abandon the snail and die, leading to further increases in the rates of parasitism of the snail hosts.
A more complex pattern emerges if we consider the community structure of parasites in two host species that share parasites. When parasites are able to use more than one species as a definitive host, their ability to establish in any one host species depends on the density of all the potential host species present in an area Because different host species may have different susceptibilities to the parasite and different parasite species may reproduce at different rates in different host species, the density of different host species will be crucial to the composition of the parasite assemblage (Dobson 1989). Variations in the population density of different host species may thus lead to variations in the parasite burdens of other host species; in some cases this may allow pathogenic parasites to establish in populations of hosts that would otherwise be too small to sustain them. Climate changes could lead to changes in the composition of host communities, which will lead to changes in the structure of the parasite community that the hosts support and the possible introduction of parasites not previously present in the host population. Where members of the parasite community are important in mediating competition between hosts, this may lead to further changes in the structure of the host community and the possible extinction of particularly susceptible hosts.
The geographical distributions of most parasite species are limited by the distributions of potential host species or by environmental constraints on the parasite's rates of development. Although developmental rates in vertebrate hosts may be comparatively unaffected by changes in environmental temperature, the available evidence suggests that the free-living stages of parasites and those that live in invertebrate poikilothermic hosts are susceptible to prevailing meteorological conditions. J. D. Gillett (1974) suggests that many vector-transmitted diseases are limited in their range because the development time of the parasite exceeds the average life expectancy of the insect vector. But increases in environmental temperature are likely to speed up development for those stages in the parasite life cycle, so long-term increases in temperature are likely to lead to increases in the ranges of many diseases transmitted by insects, such as malaria and flariasis.
Up until the mid-1970s parasitologists believed that temperature and moisture were the dominant meteorological factors determining disease outbreaks. Curiously, this area of parasitology has been relatively neglected for the last ten to fifteen years. In part, that may be because anthelmintic drugs have been developed that can be readily administered to livestock. It may also be because models for parasites now emphasize the previously neglected nonlinear components of parasite dynamics (Anderson and May 1979, May and Anderson 1979). Finally, it may also reflect the emergence of molecular immunology and the search for vaccines for parasites of domestic livestock. However, parasites are now showing serious levels of resistance to many anthelmintic drugs (Anderson and Waller 1985), and the development of vaccines is progressing more slowly than was originally anticipated. If long-term climatic changes lead to the introduction of parasites into new areas at a time when our ability to control them is rapidly diminishing, many types of domestic livestock will face major disease problems. In some cases this will lead to the abandonment of present pasture lands, which may then be set aside for nature reserves. In other regions an increasingly hungry human population will exert pressure to utilize present reserves as grazing areas. It seems unlikely that the net result of this exchange will favor wildlife.
A considerable body of literature is already available that deals with the climatic responses of a variety of parasites (Kates 1965, Levine 1963, Wilson et al. 1982). We now also have much better models for examining the dynamics of parasites at all stages of their life cycles (Anderson and May 1979, 1986; May and Anderson 1979). Although there are problems of scale associated with extrapolating between the physiological processes of parasites measured under controlled laboratory conditions and the coarser predictions available for longer-term climate change, it should be possible to merge these various sources of information to produce a quantitative synthesis of the way global climate change may affect the distribution of many parasites. It thus seems likely that global warming will give new prominence to an area of parasitology that had fallen into relative neglect.
The examples given above are mainly from well-studied species in little danger of extinction. Assessment of the potential effects of global warming on the parasites of endangered species can really only be undertaken by extrapolation from these examples and the models used to explain the more general features of parasite-host population dynamics.
A number of possible scenarios are likely to arise as host populations respond to long-term climate changes.
Consider first an endangered species whose population density has declined to such low levels that it is present only in a single nature reserve. Under these conditions it seems likely that a further decline in population size due to global warming will reduce the effects of the parasites already present in that population. However, the immigration of new host species into the area, as a response to climate change, may lead to the introduction of novel pathogens. If the endangered host has had no previous contact with these parasites, they may fail to establish, if the host is sufficiently novel, or they may establish and produce significant levels of mortality. Under these conditions, increases in the density of the immigrant hosts will lead to increases in the rates of parasite transmission. and constraints may have to be placed on interactions between the endangered species and the newly immigrating species.
Where endangered species are tolerant to increases in temperature and humidity, they are still likely to face increased assault by parasites whose transmission efficiency improves with increases in temperature and humidity (e.g., tropical diseases such as hookworm may become more important in temperate zones). Furthermore, those host populations that increase as temperatures rise are likely to suffer an increase in parasite prevalence and diversity.
If the population sizes of host species decline because of climatic changes, their rarer species of parasites and mutualists may become extinct. These species have their own intrinsic value, and they often perform a valuable function, such as the commensal chaetogasters living in the snail mantles discussed above. The absence of a parasite may be as important as its presence; some species of hosts may grow to become pests in the absence of pathogens that are now regulating their numbers.
Parasites and disease will do well on a warming earth. They are, by definition, organisms that colonize and exploit. Those species of parasite that are already common will be able to spread and perhaps colonize new susceptible hosts that may have no prior genetic resistance to them. Parasite species that are rare and have more specialized requirements may be driven to extinction. In general, these effects are likely to be worse in the temperate zone, where parasites from the tropics can colonize new hosts, than in the tropics, where parasites will have to adapt or evolve. Rare parasites that are adapted to extreme temperature, however, may become common; changes in the ranges and sizes of some host populations may allow some hitherto unimportant pathogens to become more widespread.
Aho, J. M., J. W. Camp. and G. W. Esch, 1982. Long term studies on the population biology of Diplostomum scheuringi in a thermally altered reservoir. J. Parasitol 68:695.
Anderson, R. C. 1988. Nematode transmission patterns. J. Parasitol 74:30.
Anderson, R. M., and R. M. May. 1979. Population biology of infectious diseases: Part I. Nature 280:361.
Anderson. R M., and R. M. May. 1985. Helminth infections of humans: Mathematical models, population dynamics and control. Advances in Parasitol. 24:1.
Anderson, N., and P. J. Waller. 1985. Resistance in Nematodes to Anthelmintic Drugs. Glebe, Australia: CSIRO Division of Animal Health Publications.
Armour, J., and M. Duncan. 1987. Arrested larval development in cattle nematodes. Parasitol Today 3:171.
Camp, J. W., J. M. Aho, and G. W. Esch. 1982. A long-term study on various aspects of the population biology of Ornithodiplostomum ptychocheilus in a South Carolina cooling reservoir. J. Parasitol. 68:709.
Dobson, A. P. 1986. The population dynamics of competition between parasites. Parasitology 91:317.
Dobson, A. P. 1988. The population biology of parasite-induced changes in host behavior. Quart. Rev. Biol. 63:139.
Dobson, A. P. 1990. Models for multi-species parasite-host communities. In The Structure of Parasite Communities, G. Esch, C. R. Kennedy, and J. Aho, eds., pp. 261-288. New York and London: Chapman and Hall.
Esch, G. W., J. W. Gibbons, and J. E. Bourque. 1975. An analysis of the relationship between stress and parasitism. Am. Midl. Nat. 93:339.
Evans, N. A. 1985. The influence of environmental temperature upon transmission of the cercariae of Echinostoma liei (Digenea: Echinostimatidae). Parasitology 90:269.
Ford, J. 1971. The Role of the Trypanosomiases in African Ecology A Study of the Tsetse Fly Problem. Oxford: Clarendon Press.
Gillett, J. D. 1974. Direct and indirect influences of temperature on the transmission of parasites from insects to man. Symp. British Soc Parasitol. 12:79.
Gordon. H. M. 1948. The epidemiology of parasitic diseases, with special reference to studies with nematode parasites of sheep. Aust. Vet. J. 24:17.
Gordon, R. M., T. H. Davey, and H. Peaston. 1934. The transmission of human bilharziasis in Sierra Leone, with an account of the life cycle of the schstosomes concerned, S. mansoni and S. haematobium. Ann. Trop. Med. Parastol. 28:323.
Kates, K.C. 1965. Ecological aspects of helminth transmission in domesticated animals. Am. Zool. 5:95.
Levine, N.D. 1963. Weather, climate and the bionomics of ruminant nematode larvae. Advances in Vet. Sci. 8:215.
Mackiewicz, J.S. 1988. Cestode transmission patterns. ĘJ. Parasitol. 74:60.
May, R.M. 1988. How many species are there on earth? Science 241:1441.
May, R.M., and R.M. Anderson. 1979. The population biology of infectious diseases: II. Nature 280:455.
Milligan, P.J.M., and R.D. Baker. 1988. A model for tsetse-transmitted animal trypanosomiasis. Parasitology 96:211.
Molyneux, D.H. 1982. Trypanosomes, trypanosomiasis and tsetse control: Impact on wildlife and its conservation. Symp. Zool. Soc. Land. 50:29.
Ollerenshaw, C.B. 1974. Forecasting liver fluke disease. Symp. British Soc. Parasitol. 12:33.
Rogers, D.J. 1979. Tsetse population dynamics and distribution: A new analytical approach. J. Anim. Ecol. 48:825.
Rogers, D.J. 1988. A general model for the African trypanosomiasis. Parasitology 97:193.
Rogers, D.J., and S.E. Randolph. 1986. Distribution and abundance of tsetse flies (Glossina spp.). J. Anim. Ecol. 55:1007.
Rogers, D.J., and S.E. Randolph. 1988. Tsetse flies in Africa: Bane or boon? Conserv. Biol. 2:57.
Sankurathi, C.S., and J.C. Holmes. 1976a. Effects of thermal effluents on the population dynamics of Physa Gyrina Say (Mollusca: Gastropoda) at Lake Wabamun, Alberta, Can. J. Zool. 54:582.
Sankurathi, C.S., and J.C. Holmes. 1976b. Effects of thermal effluents on parasites and commensals of Physa gyrina Say (Mollusca: Gastropoda) and their interactions at Lake Wabamun, Alberta. Can. J. Zool. 54:1742.
Shoop, W.L. 1988. Trematode transmission patterns. J. Parasitol. 74:46.
Toft, C.A. 1986. Parasites etc. In Community Ecology, J.N. Diamond and T.J. Case, eds., pp. 445-463. New York: Harper and Row.
Wallace, H.R. 1961. The bionomics of the free-living stages of zoo-parasitic nematodes: A critical survey. Helminth. Abstr. 30:1.
Wilson, R.A., G. Smith, and M.R. Thomas. 1982. Fascioliasis. In The Population Dynamics of Infectious Disease: Theory and Applications, R.N. Anderson, ed., pp. 262-319. London: Chapman and Hall.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}