 Reproduced, with permission, from:
Eldridge, B. F. 1987. Strategies for surveillance, prevention, and
control of arbovirus diseases in western North America. American
Journal of Tropical Medicine and Hygiene 37 (3): 77S-86S.
Reproduced, with permission, from:
Eldridge, B. F. 1987. Strategies for surveillance, prevention, and
control of arbovirus diseases in western North America. American
Journal of Tropical Medicine and Hygiene 37 (3): 77S-86S.
BRUCE F. ELDRIDGE
Department of Entomology, University of California, Davis, California 95616
Abstract. Present strategies for surveillance, prevention, and control of arbovirus diseases in western North America have been developed from more than 4 decades of epidemiological research and development of mosquito control technology. Methods of prediction of outbreaks remain imprecise, although our understanding of sources of variation associated with indicators used for prediction is improving. Well organized and funded systematic mosquito abatement remains the most effective method of prevention of human cases of mosquito-borne virus disease, although emergency methods must be employed when outbreaks are imminent. The development of information management systems technology, use of recent developments of sampling theory, and research on vector competency and related areas should permit much better precision in estimates of impending outbreaks.
The development of strategies for the surveillance, prevention, and control of arthropod-borne virus diseases in North America grew out of studies of the epidemiology of these diseases conducted during the period preceding World War II. Approaches have been refined over the years, and materials and equipment have been improved, but the basic concepts remain relatively unchanged since those times. I will examine these concepts and discuss their validity in light of the present state of knowledge of arbovirus disease ecology. If the name "Reeves" appears frequently in this discussion it is because his contributions to this area have been so extensive.
Early concepts
The objective of arbovirus disease surveillance is to assess the existing epidemiological situation for a disease so as to be able to predict the likelihood of human disease outbreaks, and thus permit intervention to avoid or abort such outbreaks. This objective depends upon the concept that various phenomena such as high mosquito population densities precede human disease cases, and that these phenomena can be used as indicators to predict disease outbreaks. The use of these indicators for this purpose relies on an understanding of the basic components of the disease cycle: pathogen, vectors, and hosts. Thus incrimination of vectors and pathogen, and identification of hosts must always receive the highest priority in epidemiological studies of newly emerging diseases. In the case of western equine encephalomyelitis (WEE) and St. Louis encephalitis (SLE) in western North America, it was only after Culex tarsalis had been incriminated as the primary vector, and birds as the primary hosts, that sound surveillance methods could be developed. Studies which resulted in this incrimination were described in a series of papers published by Hammon, Reeves, and associates in 1941 and 1942.[1-8] These papers are significant not only because they established the basic ecological pattern of WEE and SLE in western North America, but also because of their contribution to the concept of vector incrimination and host identification. Although physiological and ecological factors underlying concepts of vector relationships in arboviruses have been explored and elucidated over the years,[9,10] the fundamental approaches of vector and host identification remain those used in the classic studies in the Yakima Valley in the early 1940s. Four criteria formed the basis for incrimination of Cx. tarsalis as a vector of SLE and WEE viruses.[5,6] Paraphrased from the explicit definition of Barnett,[11] incrimination of an arthropod species as a vector of a given pathogen requires demonstration that the species feeds upon humans under natural conditions, demonstration of a biological association in time and space with the occurrence of clinical or subclinical human infections, the repeated isolation of the completely identified infectious stage of the pathogen, and demonstration under controlled conditions of efficient transmission of the pathogen by the arthropod species to a suitable host.
Research conducted since these criteria were established has not invalidated them, although subsequent studies have occasionally ignored them. But the demonstration that geographic strains of Cx. tarsalis can vary in their susceptibility to WEE virus infection[10] and the natural occurrence of WEE virus strains with altered host virulence[12] emphasize that results from one area or time may not necessarily apply to another. This may restrict the applicability of surveillance techniques, and necessitate broad geographical coverage for maximum reliability. This is not a new concept. The need to determine vector status for mosquito species separately for each geographical area was stressed earlier by Reeves[13] in his outline of ecological determinants for arbovirus disease outbreaks.
Contemporary elements of surveillance
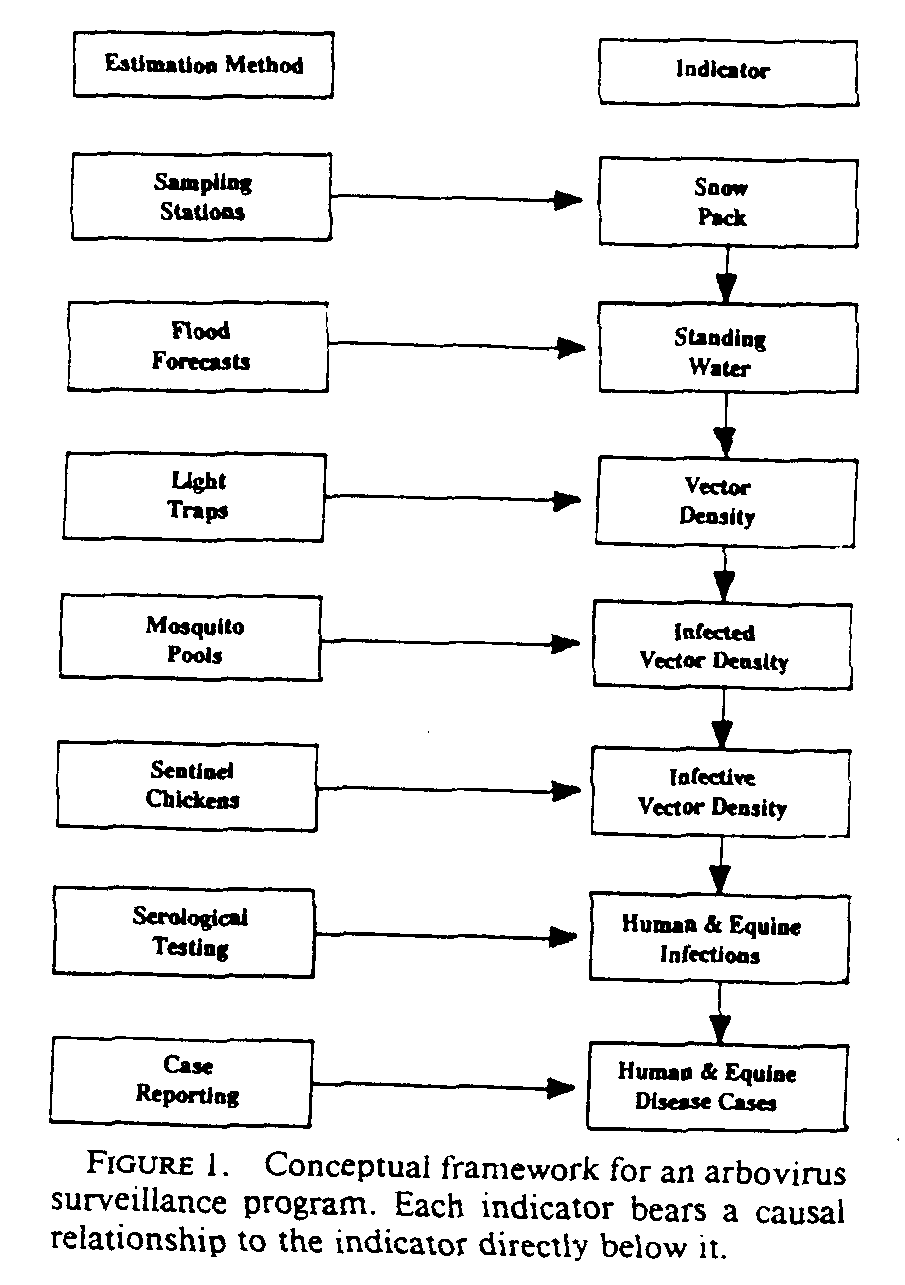
The California Mosquito-Borne Encephalitis Virus Surveillance and Control Program[14] consists of environmental and biological components. Environmental factors include monitoring of snowpack in mountains, spring rainfall, flood control reservoir releases, and seasonal temperature patterns. Biological components include sampling of mosquitoes to estimate population levels, testing of mosquito pools to estimate infection rates, periodic bleeding of sentinel chicken flocks to detect seroconversions, and monitoring of human and equine cases of disease. Reeves and Milby[15] discussed the relative value of these components and suggested the addition of another: economic resources available for mosquito abatement. They showed a strong correlation between funds available for mosquito abatement in California and WEE seroconversion rates in sentinel chickens for the year preceding, and the 2 years following passage of a property tax limitation measure in 1978.
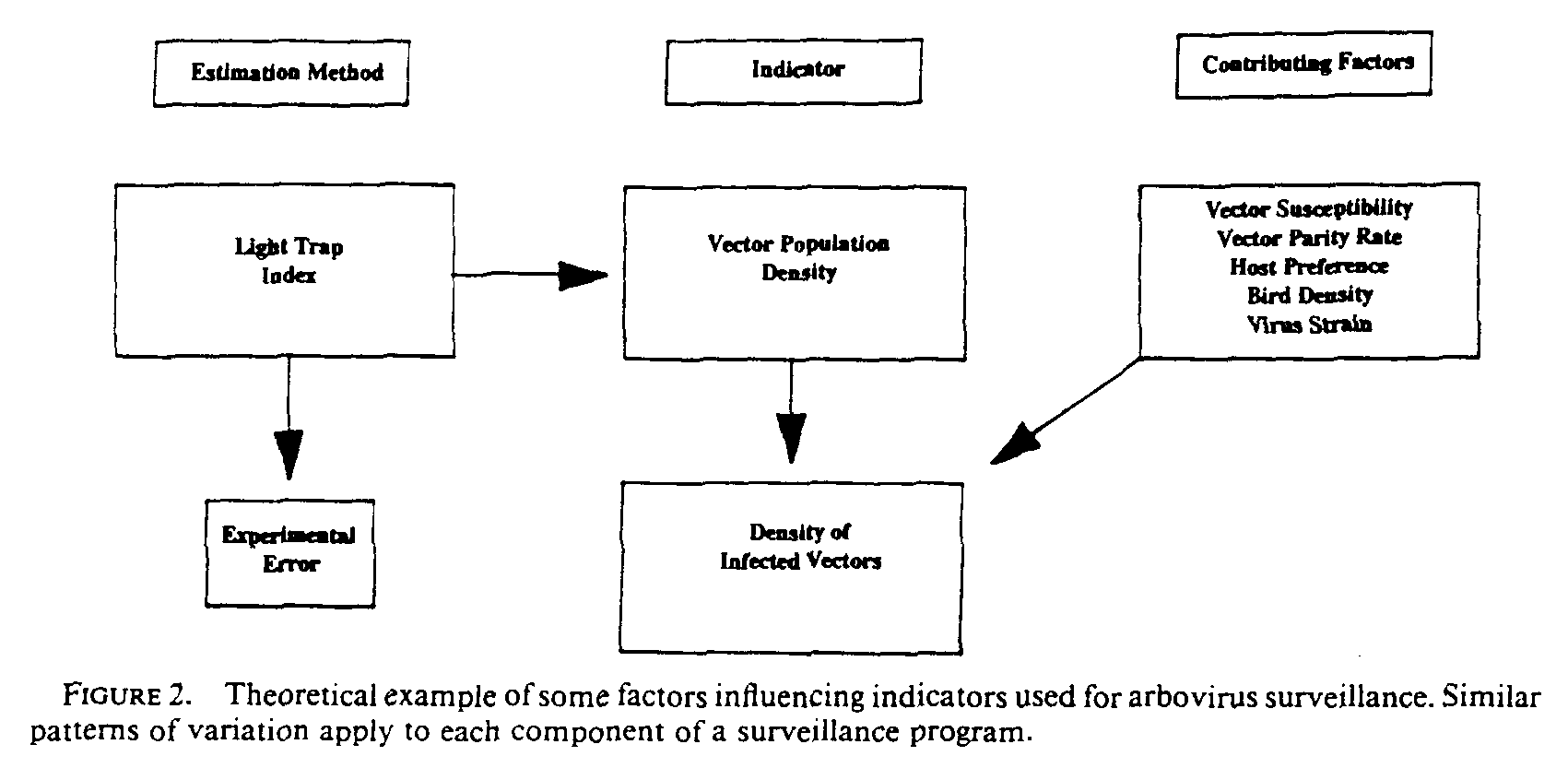
Surveillance programs must combine a number of components in order to produce accurate forecasts of human disease activity (Fig. 1). The various components are subject to different sources of variation and therefore differ in their sensitivity as predictors, and in the amount of time they provide in advance of human infections. There are 2 types of errors which can result in a lack of correlation between the measurement of various factors and cases of human disease. One type results from the failure of sampling data to estimate accurately the true value of the factor, the other from the degree to which the factor being estimated departs from an actual biological relationship with human disease cases. There are causal relationships among the various indicators, and for any given indicator, there will be other contributing factors which will affect it (Fig. 2).
Analysis of weather data and groundwater. Two major environmental factors are related to arbovirus disease outbreaks: temperature and precipitation. These 2 factors control another factor: available water for mosquito breeding. Later in the year, temperature is related to rate of development of immature stages of mosquitoes and to virus replication and transmission rates. There are yet other temperature-dependent biological phenomena which influence virus activity, such as diapause, autogeny, and virus strain selection.
Hess et al.[16] examined the relationship between temperature and outbreaks of WEE and SLE in North America and concluded that outbreaks of WEE occurred at or above the 70deg.F June isotherm, outbreaks of SLE below it. They further concluded that enzootic activity could be predicted by observing spring temperature accumulations. Years in which spring temperatures were high (accumulation of 10 day-degrees above 75deg.F before the end of spring) were favorable for SLE outbreaks, but not WEE. Years in which temperatures were low (10 day-degrees above 70deg.F not accumulated until early summer) were favorable for WEE virus activity, but not SLE. Reeves and Hammon[11] also noted the relationship between high temperatures and SLE outbreaks, but added the importance of heavy winter snowpack followed by heavy spring run-off. For a given year, depth of snowpack in the western United States is the earliest factor which can be evaluated in relationship to probability of disease outbreaks, but it is also the most remotely related, and thus the least sensitive.[15] Later in the year, spring run-off, river flows, water tables, and other measurements of groundwater can be studied.
Retrospective studies of WEE and SLE epidemics in California have shown that some have been associated with abnormally large winter snowpack and spring flooding (e.g., 1952), but that these conditions have not invariably led to epidemics.[13, 18] The most direct result of excessive amounts of water present during spring would be an increase in the size of the population of vector mosquitoes. If mosquito abatement agencies are alerted early to the probabilities of large populations of vectors, however, abatement efforts can be intensified, and the build-up of the vector population aborted. This may have occurred in California in 1958.[18]
Weather and hydrologic data are now commonly available on computerized data bases. This would seem to be an area where modern technology could be used to considerable advantage. Computer-based geographic information systems[19] have the ability to provide highly realistic simulations of flooding for specific areas based on inputs such as precipitation and snowpack. However, although the concepts are technologically feasible, implementation of practical systems are a long way off. This situation may continue until the technologies of geographic information systems and remote sensing systems are integrated.[20]
Sampling of vector mosquitoes. Sampling for arbovirus surveillance is done on the assumption that there is a positive correlation between vector density and incidence of human disease.[21, 22] Mosquito population estimates also are useful in assessing the success of control operations. In North America, the Standard New Jersey light trap,[23] or some modification of it, sometimes called the American New Jersey light trap,[24] is used almost exclusively for routine surveillance. There are several disadvantages to these traps, most notably their bulkiness and need for a source of 120 or 240 volt alternating current (although battery-powered models are available), but their widespread acceptance as the standard for routine surveillance assures their continued use. There are literally decades of mosquito collection records available throughout North America based on these traps.
Longshore et al.[25] reported the lack of a correlation between monthly light trap indices with cases of WEE and SLE in California statewide, as well as in Kern and Fresno counties, for the years 1953-1957. Blackmore et al[21] also were unable to show a direct correlation between densities of Cx. tarsalis and virus transmission rates to sentinel birds in Colorado over a 3-year period. Reeves,[26, 27] on the other hand. working with data from Kern County, California, found a positive correlation between season-long light trap indices for Cx. tarsalis and both SLE and WEE. Based on these studies, Reeves suggested that if the density of Cx. tarsalis could be lowered to less than 10 females per trap night, SLE virus would disappear from the environment. and below 1 female per trap night, WEE virus would disappear. Reeves[27] pointed out that densities of Cx. tarsalis were often several times higher in the Sacramento Valley than in the southern San Joaquin Valley (often > 500 Cx. tarsalis per trap night in the former) but without evidence of high WEE or SLE virus infection rates and also that, in general, extremely high densities of vectors appear to be associated with low efficiency of transmission. Olson et al.[28] compared New Jersey light trap indices for a number of areas throughout California with mosquito infection rates, sentinel chicken seroconversion rates, and human cases of WEE and SLE over the 21-year period 1953-1973, and demonstrated a positive correlation between the indices and all 3 factors. They also confirmed the earlier finding of Reeves[26] that at light trap indices above these levels, incidence of both diseases in humans was lower, falling to zero at the highest indices. In rural areas, peaks in human cases corresponded to light trap indices several times higher.
The reasons for lower virus activity levels associated with very high light trap indices are not fully known. Reeves[9, 26] suggested that at very high mosquito densities, birds exhibit mosquito avoidance behavior, causing a shift to mammal feeding and thus resulting in low virus transmission efficiency. Reeves et al.[29] showed that as the number of female Cx. tarsalis captured in chicken-baited traps increased, the proportion which had bloodfed decreased, but if the hosts were restrained nearly all female mosquitoes took a bloodmeal. Shifts in feeding by Culex mosquitoes from birds to mammals during summer have been demonstrated[30-32] and Edman et al.[33] also concluded that host defensive behavior associated with high vector densities cause these shifts. Edman et al.[33] have emphasized the complexity of phenomena associated with success of bloodfeeding behavior and point out that factors in addition to host defensive behavior are also involved. Analysis of the parity status of light-trapped females at various densities would be helpful. If there were a disproportionate number of nulliparas present in large trap samples, the appearance of increased transmission efficiency could result.
Sources of experimental error in light trap data are well known to all who have operated these devices. Service[24] extensively discusses reasons for variation in results. Even in records based on a single type of trap, such as the "Standard" New Jersey trap, considerable trap-to-trap variation is present because of differences in trap design, as well as because of variation in motor and fan efficiency.[34] Light traps do not sample all mosquito species in proportion to their actual numbers present. This may be because of differences in attraction to different species, and also to differences in spatial distribution of different species. Although light traps are sometimes regarded as sampling devices for the estimation of mosquito population density, they are biased toward host-seeking females, and consequently estimate the density of only a subpopulation. This may be quite satisfactory for the purpose of predicting the probability of virus transmission, if the sampling of nongravid blood-seeking females is itself unbiased. Trap placement is critical in this regard, and location of traps close to larval breeding sources, or close to sources of bloodmeals must be avoided. Unfortunately, it is seldom possible to determine trap placement after the fact.
Although cases of human disease presumably will not occur if density is below certain levels, high densities are not invariably associated with human disease. Some of the reasons for this have been discussed. Population density estimates obtained from light trap data do not differentiate between parous and nulliparous mosquitoes, and there is no way from inspection of light trap data to know anything about the vector competence the population being sampled. Differences in virus strain infectivity for mosquitoes, low virus susceptibility of mosquitoes, low population levels of birds, altered host contact, and lack of a virus source are all factors which can cause low correlations between vector population density and vector infection rates.
Virus isolations from mosquito pools. The routine testing of mosquito pools for isolation of virus can provide valuable epidemiological information about the current status of arbovirus diseases. Such information has not been regarded to be a sensitive predictor of human disease.[35] but theoretically, should be more sensitive than estimates of overall vector density. In some studies, SLE virus infection rates in mosquitoes over several years have not shown a high positive correlation with cases of human disease.[35] Kokernot et al.,[36] however, did find a positive correlation between minimum infection rates of mosquitoes (primarily Cx. pipiens) and human cases of SLE in studies conducted over several years after an epidemic of SLE in Illinois. There are many reasons why virus infection rates in mosquitoes may not show a high positive correlation with cases of human disease. As emphasized by Reeves et al.,[18] high vector infection rates are not necessarily followed by high transmission rates. In field studies, Reeves et al.[29] showed that infection rates of chickens exposed in can-traps were always less than infection rates in mosquitoes trapped in the can-traps. They reasoned that for any given group of infected mosquitoes, some proportion will not have been infected long enough for the virus to have completed its extrinsic incubation period, and those mosquitoes will therefore not be infective. Hardy et al.[10] suggest that additional factors are at work as well. They suggest that female Cx. tarsalis can modulate infections of WEE virus so that some proportion of infected females never undergo infection of salivary glands. Kramer et al.[37] found that in laboratory experiments, among female Cx. tarsalis which were infected at relatively low doses of WEE, 38% were unable to transmit virus orally. Hardy et al.[10] refer to this as the "salivary gland infection" barrier. Differences in susceptibility to WEE virus infection among naturally occurring populations of Cx. tarsalis, have been reported,[35] but not differences in transmission rates among populations equally susceptible to infection. However such differences have been observed in laboratory strains.[37]
There are other possible reasons for lack of agreement between estimates of mosquito infection and transmission rates. Estimates obtained by detection of virus in pooled mosquitoes may not accurately represent actual infection rates in the mosquito population being sampled. Chiang and Reeves[39] provided an excellent guide to the statistical estimation of virus infection rates in mosquitoes. They point out that confidence limits for estimates based on testing of pools of mosquitoes can be very broad when a high proportion of pools are positive for virus, especially when relatively few pools are available for testing. Further, it has been shown that suspensions of normal adult mosquito tissue may interfere with detection of small amounts of virus in cell culture systems.[40] There may be ecological factors involved as well. Mosquito collection sites may not be sufficiently close to locations where human infections are taking place to represent samples of the population of mosquitoes transmitting virus. The species of mosquito yielding the greatest number of virus isolations may not necessarily be the primary human vector, although this has not been shown to be the case in the western United States. It is certainly true for eastern equine encephalitis (EEE) in the eastern United States, however, where Culiseta melanura consistently is found infected in large numbers, yet seldom feeds on human hosts.
Improvements in virus isolation techniques seem likely in the near future. Tests such as the enzyme immunoassay[41, 42] have been shown to be rapid and sensitive for virus isolation. Until such tests have been extensively field-tested, however, conventional methods such as mouse inoculation will continue to be used.
Sentinel chicken flocks. Domestic fowl, especially chickens, have been widely used for routine surveillance of arbovirus activity. The suitability of chickens as WEE and SLE sentinels was demonstrated by showing that they became infected in nature and that experimentally, after inoculation with relatively small doses of virus, became infected[43, 44] and circulated hemaggluttination-inhibition antibodies at detectable levels for at least a year.[45] If the chickens are bled frequently, seroconversion can be used to indicate time of transmission of virus, and seroconversion rates of chicken flocks collectively will indicate overall virus activity. Since seroconversion rates in sentinel chickens are related proportionally to numbers of infective vectors present, they should be sensitive predictors of human infections. Generally, seroconversion rates for SLE virus in sentinel chickens have correlated well with human cases.[35] One rationale for using chicken flocks as sentinels in addition to testing of mosquito pools for virus is that seroconversions of sentinel chickens provide an indication of virus transmission, whereas positive mosquito pools indicate only infection of mosquitoes. Chickens are not useful as sentinels for all situations. Crans[46] reported that they did not serve as early warning vehicles for an EEE epizootic in New Jersey in 1984, although they did exhibit seroconversion late in the year. In the 1975 outbreak of WEE in Manitoba. however, the first sentinel chicken seroconversion occurred approximately 1 month before the first equine case. and at least 2 months before the first human case.[47] Sentinel chickens are probably most valuable when levels of virus activity are low.[15] When minimum infection rates in mosquitoes are below 1:500, it is impractical to collect and process enough female mosquitoes to obtain reliable estimates of mosquito infections. However, seroconversions of sentinel chickens for WEE virus can be observed at these times.[15] Disadvantages of chickens as sentinels include the lag between infection and development of antibodies, the labor involved in the care and bleeding of individual birds, and the possibility of nonspecific antibody reactions.[15] The ability of sentinel flocks of chickens to serve as early warning devices is probably most dependent upon proper placement of flocks and on frequent bleedings early in the season.
Surveillance of horse cases. In the case of WEE virus, which can produce serious disease in horses, the recording and analysis of cases of equine disease can be useful as a surveillance tool if reporting is thorough and diagnostic facilities are available. Although widespread vaccination of horses has reduced the incidence of WEE in these animals in the western United States, horse cases continue to occur sporadically, especially in years of high virus activity. Unfortunately, not all states have a diagnostic capability for WEE virus, and must depend on federal laboratory support. In some states, cases of WEE in horses are not systematically recorded and reported. Past records of confirmed equine WEE cases probably represent only a fraction of the actual number of cases, and attempts to correlate human and equine disease are not rewarding. There are also epidemiological reasons for a lack of correlation between cases of disease in the 2 hosts. There may be divergence over time in levels of exposure[48] and in immunity status. Finally, other mosquito species such as Aedes melanimon may be involved in the transmission of WEE virus to horses.[49]
Human disease surveillance. A key element in surveillance is the diagnosis and follow-up of human disease cases. The procedures involved have been well described by Bowen and Francy.[35] Although the season-long pattern of human cases may have little predictive use for the year in which they occur, cases early in the year can alert public health officials for the need to marshal resources for emergency disease control procedures. Also. the occurrence of human cases one year should result in better readiness for the following year. Since, as indicated above. surveillance activities must be relatively specific in terms of geography, occurrence of human cases can help in choosing strategies for geographical coverage.
An abundance of information is available on specific tactics available for the control of arbovirus diseases, most of it devoted to mosquito control. A discussion of the details of these tactics is beyond the scope of this paper. Since vector control is presently the only practical approach to control of the mosquito-borne encephalitides, I will ignore other subjects, such as vaccine use. From the standpoint of strategy, options for vector control may be divided into 2 classifications: normal procedures carried out to control mosquitoes regardless of the level of virus activity, and emergency procedures carried out when the threat of human virus disease is high, or actually occurring. Some reaction plans, such as that of the state of California, divide the second classification into 2 readiness levels: one based on early indications of the likelihood of cases, the other based on an actual outbreak.[14]
A persuasive argument can be made that one of the factors which has led to the gradual reduction of human cases of WEE and SLE in California over the past 2 decades has been the development of organized mosquito abatement practices. This is one of the conclusions reached by Reeves et al.[18] in their analysis of the factors associated with the lack of human of cases of WEE in 1958, a year in which a number of factors indicated a high probability of an outbreak. A key element in this argument is acceptance of the principal that outbreaks of disease in humans and horses are associated with unusually high densities of vector mosquitoes, and that virus transmission will not be maintained when vector density falls below some minimum level. The correlation between Cx. tarsalis density and human cases of WEE and SLE over a 21-year period seems well established[28] and although the minimum density of Cx. tarsalis needed to sustain virus transmission is extremely difficult to define precisely, the levels recommended by Reeves[26, 27] are reasonable goals, after adjustments are made for geographic location and ecological situation.
To have an impact on prevention of encephalitis in humans, normal mosquito abatement activities must stress control of the primary vectors. Mosquito larviciding and source reduction form the basis of these activities. Problems in physiological resistance to conventional pesticides, especially organophosphates,[50] have hampered these efforts. but newer materials such as Bacillus thuringiensis serotype H-14 (Bti) and larvicidal oils have continued to provide effective control of larvae.
Emergency control measures are predicated on the assumption that infective adult mosquitoes are present in large numbers and must be eliminated in the shortest time possible.[51] The constant monitoring of insecticide susceptibility in populations of the primary vector species is a vital prerequisite to emergency control measures. There is rarely time to conduct such testing after an outbreak has developed or is imminent, and there have been several studies which have demonstrated nearly complete lack of control when resistance has been a factor.[52] Current practice for emergency control of infected vectors is the aerial application of insecticides using the ultra-low volume (ULV) technique.[53, 54] In theory, treatment should result in the elimination of infected, parous females so that even if the population recovers in a week or so, parity rates should be much lower, and the virus transmission rate should be greatly reduced. This assumes that recruitment is primarily from newly emerging adults rather than from immigrants from untreated areas. The success of this approach in achieving a rapid elimination of virus transmission has not lived up to expectations in every instance. There are reports of large decreases in the density of mosquitoes following treatment,[55] and of decreases in virus infection rates in vector mosquitoes,[56] but in most instances where natural populations have been examined critically before and after treatment, the reduction of virus infection rates in mosquitoes and vertebrate animals has been difficult to document. Nearly all studies have shown that post-treatment reductions have been transitory. Most authors have concluded that infiltration of adults from untreated areas have caused population indices to quickly rise to pretreatment levels.[57] When this happens, the overall parity rate and the infection rate of adult mosquitoes is not greatly affected. Thus Mitchell et al[58, 59] found that control of Cx. tarsalis in Hale County, Texas, lasted for only 3 days after treatment with ULV malathion from aircraft, and failed to demonstrate a significant reduction in virus or wild bird virus infection rates, or in subclinical infection rates in children. Brust and Ellis[55] reported similar results in Manitoba during the 1975 epidemic of WEE. They estimated that after insecticidal treatment with Abatereg. which resulted in the destruction of about 80% of Cx. tarsalis adults present, the population remained suppressed for only 3-5 days. Reisen et al.[60] studied the effect of ULV aerial application of chlorpynfos and resmethrin on the age structure of Cx. tarsalis populations in Kern County, California. They reported only temporary suppression of adult populations, but did find that the minimum infection rate of WEE virus in Cx. tarsalis declined after resmethrin treatment of one of the study areas. Reisen et al.[61] obtained mixed results in assessing parity status and mosquito infection rates (WEE) after spraying with propoxur from aircraft in Kern County, California. In some sites, they were able to show alteration of parity status and infection rates, but in other sites, the minimum infection rates remained at greater than 1 in 1,000 in the sprayed area. McHugh and Washino[62] reported short-term control of Cx. tarsalis with malathion applied from helicopters and found no differences in parity rates in populations from treated and untreated areas. They concluded that immigration of mosquitoes from untreated areas and protection of resting adults by vegetation prevented long-term control. Two major differences mark the early and reportedly successful attempts at epidemic control of vectors by aerial ULV treatment and later controlled experiments to prove the effectiveness of such treatments. The early attempts involved very large treated areas (3.2 million acres in the case of the spraying of the Gulf Coast following Hurricane Betsy in 1967[63]) and were aimed at vector populations that apparently were susceptible to malathion. Neither situation pertained to the California tests described above.[52, 61, 62]
There are a number of factors which affect the efficacy of adulticiding as an emergency control measure. Less than ideal weather factors, equipment limitations, vegetative cover, poor timing, insecticide resistance, and improper application technique can all contribute to poor control. Proper selection of equipment is very important.[64] Proper timing of sprays is dependent upon the flight habits of the target mosquito species, which is not known. Reisen et al.[61] concluded that late afternoon spraying was more effective than morning spraying for control of adult Cx. tarsalis in Kern County because of greater synchronization with the diel activity cycle of adults. in spite of conditions traditionally considered less than ideal for spraying. Even under ideal conditions, repeated adulticide treatments are probably necessary to lower adult population levels sufficiently to stop transmission, especially if the treatments are done early in the season. Reisen et al.[61] recommended 3 treatments at 3-day intervals.
The many variables which must be considered in conducting emergency adulticiding, and the studies which have demonstrated that it is very difficult to lower density of vectors to levels below that needed to stop transmissions emphasize that such spraying should be regarded as a remedy to be used only after all preventive measures have failed. Well organized mosquito abatement efforts which include adequate routine sampling for density, vector competence, virus infection, and insecticide susceptibility, are far more effective and, in the long run, more economical.
24. Service, M. W., 1976. Mosquito Ecology, Field Sampling Methods. Halstead Press, New York.
{kind=link}
{kind=link}