 Reproduced, with permission, from:
Patz, J. A., T. A. Burke, and P. R. Epstein. n.d. Public health effects of global climate change and ozone depletion: Implications for North America. Unpublished draft.
Reproduced, with permission, from:
Patz, J. A., T. A. Burke, and P. R. Epstein. n.d. Public health effects of global climate change and ozone depletion: Implications for North America. Unpublished draft.
Jonathan A. Patz, MD, MPH *
Department of Environmental Health Sciences
Johns Hopkins School of Hygiene and Public Health
615 N. Wolfe St., Room 7041
Baltimore, MD 21205-2179
phone: (410) 955-4130 (work)
(410) 377-5623 (home)
FAX: (410) 955-1811
Thomas A. Burke, PhD, MPH
Department of Health Policy
Johns Hopkins School of Hygiene and Public Health
Paul R. Epstein, MD, MPH
Department of Medicine
The Cambridge Hospital
Harvard Medical School
* Send Correspondence
Objective.-- To assess public health consequences of global warming and stratospheric ozone depletion likely to impact North America.
Data Sources.-- Data were obtained from articles accessed through the National Library of Medicine's MEDLINE database and from personal communication with key investigators.
Study Selection.-- Studies relevant to health effects of global warming or ozone depletion were reviewed. We concentrated on UV-induced immunosuppression, vector-borne disease in North America, and the public health implications of changes in marine ecology.
Data Extraction.-- Guidelines for assessing data quality included study design, biologically plausible outcomes, dose-response relationships, and consistency with similar work.
Data Synthesis.-- Review of the existing literature suggested that in the United States, mosquito-borne arboviruses are likely to extend northward due to warming trends and altered patterns of precipitation. Dengue Fever and Saint Louis Encephalitis, against which no vaccines or antiviral agents have been developed, may pose a significant risk. Cholera and "red tides" may continue to threaten coastal populations as phytoplankton blooms proliferate in warmer, nutrient-rich waters. Increased UV-B radiation contributes to cataracts, skin cancer, and of particular focus for this section of the review, immunosuppression.
Conclusions. -- Primary prevention of global climate change through environmental protection ranks as the top priority. To the extent, however, that some climate change may be unavoidable or is already occurring, secondary prevention is warranted. Recommendations include internationally coordinated monitoring for the emergence of epidemics, laboratory and field studies of vector ecology, and elaboration of UV-B effects on the human immune system.
KEY WORDS: arboviruses, cataract, cholera, climate, encephalitis, immunosuppression, mosquitoes, public health, skin neoplasms, ultraviolet rays
(Non Index Medicus key words: global warming, ozone depletion)
INTRODUCTION
The United Nations Conference on the Environment and Development (UNCED) in Rio de Janeiro last June brought world focus to the problems of global warming and ozone depletion. Energy sources, rainforests, species biodiversity, and economic development received the bulk of attention. While there was much concern about the indirect effects on populations predicted from meteorologic general circulation models, i.e., flooding due to thermoexpansion of oceans[1,2] food shortages from crop damage[3], diminished fresh water supply[1], and climate-induced migration[4], analysis of the more direct health outcomes of global climate change was all but absent from the official agenda[5]. This paper examines some of the existing and potential public health consequences which may specifically threaten temperate regions and makes recommendations for prevention. Debate over meteorologic predictions is beyond the scope of this review.
Carbon dioxide, the most abundant greenhouse gas, has increased by 25% since the preindustrial era[1]. A 1-5deg. C rise in temperature is the forecasted greenhouse effect if preindustrial carbon dioxide levels double in the next quarter century[6]. Plants may initially benefit from higher CO2 and temperatures which allow more efficient water use during photosynthesis[7]. However, crop damage from UV-radiation and reduced rainfall in mid-continental regions may offset potential gains[3]. Reviews addressing the medical consequences of global warming have described a broad array of potential outcomes, from increased heat stroke and asthma, to altered distribution of vector-borne diseases and cholera[3,4,8-11].
Stratospheric ozone depletion specifically allows penetration of UV-B (290-320 nanometers), the most biologically destructive wavelengths[12,13]. UV-B levels are expected to rise disproportionately in temperate and arctic zones[14], since a clear relationship has been established between higher latitudes and the extent of ozone thinning[15]. The U.S. Environmental Protection Agency (EPA) estimates an average 2% rise in UV-B per 1% drop in stratospheric ozone[16], increasing the risk of cataracts, skin cancers, and immunosuppression. (Table 1).
GLOBAL WARMING AND PUBLIC HEALTH
Heat Related Illness
Temperate and polar regions are expected to warm disproportionately more than tropical and subtropical zones[6]. Kalkstein[17] has determined that key meteorologic variables contribute to heat stroke incidence, the most significant being "oppressive" nighttime temperatures; the greenhouse effect specifically would create these conditions[18]. A greater than 6-fold increase in heat-related deaths could result in the U.S. from the expected temperature change of doubling atmospheric CO2 (though acclimatization may dampen the impact)[13,19].
In a warmer world, respiratory illness will be exacerbated since higher temperatures accelerate the chemical reactions which catalyze the formation of acid aerosols[20]. Tropospheric ozone production from increased UV radiation's photochemical reaction with volatile organic compounds and nitrous oxides in urban smog will compound respiratory illness; ozone is a potent pulmonary irritant[21].
Vector-borne infectious diseases
Vector-borne diseases in temperate regions may become difficult to control as insects penetrate higher latitudes. and as warmer temperatures shorten incubation periods of vectors and pathogens[22]. The spread of insecticide and drug resistant organisms may further handicap efforts to control epidemics. Conversely, reductions in some diseases in specific regions may occur; Rocky Mountain Spotted Fever, for example, may decline due to ticks' intolerance of high temperatures and diminished humidity in the South[23].
Geographic range and vector infectivity rate are the two most important determinants of vector transmission of disease[24]. These are not only affected by rainfall, temperature and humidity, but by vegetation, insect predators, and competitors as well. Human activity and proximity to stagnant water (ponds, marshes, tire dumps, tanks etc.) are the crucial elements for maintenance of infection in a human population. Shope[22] emphasizes the importance of assessing the following factors in predicting the effect of global climate change on the distribution of vector-borne diseases: 1) the current geographic distribution of the disease; 2) the range of the non-human reservoirs (i.e., insect or mammal); 3) temperature related arthropod vector development and pathogen replication; and 4) reservoir and pathogen adaptation, or capacity for migration. Human cases of mosquito-borne diseases are a result of leaks in the enzootic cycle, where disease escapes from the normal passage between vector and non-human host or reservoirs[25].
Temperature determines vector infectivity by affecting pathogen replication, maturation, and the period of infectivity[24]. Snails carrying schistosomiasis, for example, become non-infectious as water temperature cools in fall[3]. Elevated temperature and humidity also intensify the biting behavior of most insects and are associated with increased human host vulnerability through removal of protective clothing.
According to the EPA, mosquito transmitted diseases such as malaria, yellow fever, dengue, and arboviral encephalitis pose the greatest vector-borne threat to the United States if average temperatures rise[24]. Malaria ranks first in morbidity and mortality worldwide[4], but prophylactic and definitive pharmacotherapy are available to combat this illness. Unfortunately, drug resistant strains are a major problem for Plasmodium falciparum and now vivax[26], and changes in climate could alter the range of the Anopheles mosquito. Yellow fever, carrying a 20-50% mortality rate[27], may become a significant threat in the southern U.S. An effective vaccine is available, however, and could be generally administered (above 9 months of age)[28] with amplified production. Dengue fever and arboviral encephalitides are, therefore, the vector-borne diseases of greatest concern for temperate regions in the event of global warming.
Dengue Fever
According to the American Academy of Pediatrics, "The prevalence of different arboviral diseases is related to climatic conditions that affect the prevalence of the specific vector."[28] Aedes aegypti is the primary vector of dengue and yellow fever, and its range is limited by cold weather, which kills both larvae and adults. Its present northern extent in the U.S. is 35deg. latitude, corresponding to Memphis, Tennessee[27].
Dengue fever is an acute febrile illness; infection by two of the four serotypes is associated with Hemorrhagic Fever, which when treated, still carries a case fatality rate of 5%[27,29,30]. Dengue is widespread in the Caribbean, tropical America, the South Pacific, Asia, and Africa[31]. Since 1950, over 1.5 million cases have been reported from southeast Asian cities[32]. First appearance in the Western Hemisphere was documented in Cuba in 1981[33]. Global warming could extend the range of the vector, permit earlier transmission by hastening mosquito larval development, and lengthen the season in which epidemics may arise[27]. Following a mosquito blood-meal, viral replication to achieve infectivity takes 1-2 weeks[29]. Temperature rise would shorten this "extrinsic" incubation period. Cooler climes have thus far limited the persistence of imported dengue in the U.S., for Ae. aegypti does not survive freezing temperatures[27].
In 1985, the Asian tiger mosquito, Aedes albopictus, was detected in Texas[31]. Ae. albopictus, a competent vector for dengue, yellow fever, and several encephalitides, has become established in the Western Hemisphere in 21 contiguous states, Hawaii, and parts of Brazil[34]. Ae. albopictus range extends 7deg. further north than that of Ae. aegypti[31]; a suspended physiologic state, or "diapause", enables its eggs to delay hatching and withstand cold temperatures[29]. But Ae. albopictus is a ubiquitous biter, and while it may function as a bridge vector, it may not easily serve to maintain infection in the human population. The multiple meteorological and biological (including adaptive) factors involved in establishing and maintaining vector-borne diseases makes for substantial uncertainty in predictions; one can only say that distribution and impact will likely be altered.
Saint Louis Encephalitis
When non-human reservoir hosts harbor viruses within the disease transmission cycle, eradication by vaccine becomes virtually unattainable. Passerine birds serve as the principal vertebrate host of Saint Louis encephalitis (SLE)[35,36], which originates from the same virus family as dengue and yellow fever, Flaviridae. SLE is the leading cause of arboviral encephalitis in the U.S., accounting for 65% of diagnosed cases[37,38], and from 1977 to 1986, nine discrete outbreaks occurred in the U.S[39]. There are several vectors for SLE: in the rural west SLE is endemic and is transmitted by Culex tarsalis; in Florida, C. nigripalpus is the primary vector; in the urban southeast, C. quinquefasciatus, and in the urban north C. pipiens are the dominant vectors. Both urban species require water with high organic content (i.e., sewage or fertilizers)[22].
Clinical features of SLE range from headache to aseptic meningitis and encephalitis, with a case fatality rate of 20-80% among the elderly[22]. As with dengue, diagnosis is made by serum and cerebrospinal antibody titers or viral isolation[30]. Epidemics generally occur south of the 70deg. F June isotherm, but northerly outbreaks have occurred during unseasonably warm years[22]. Reeves[40] describes factors necessary for a SLE epidemic: 1) presence of virulent strains; 2) large numbers of avian hosts; 3) competent vector species; 4) a population of susceptible humans; and 5) sufficiently high environmental temperatures to insure a short extrinsic incubation period.
Human outbreaks are highly correlated with several-day periods when temperature exceeds 85deg. F[40]. In 1984, the major California epidemic followed extremely high temperatures[39]. Precipitation patterns are also important for transmission[38]. Heavy winter rainfall combined with summer droughts are characteristic of epidemic years. Low summer rainfall in urban areas resulting in less wash-out of sewage, preserves C. pipiens breeding sites[22]. Furthermore, computer analysis of monthly climate data has demonstrated that excessive rainfall in January and February, in combination with drought in July, most often precedes SLE outbreaks[41]. Such a pattern closely resembles computer predictions for climate change in temperate zones associated with global warming: warm, wet winters and hot, dry summers[6].
OCEANOGRAPHIC CHANGES AND PUBLIC HEALTH
Oceans comprise 70% of the earth's surface and play an important role in climate modulation. Conversely, environmental variations impact marine population dynamics in the lowest trophic level of earth's oldest food chain: phytoplankton (algae) and zooplankton. Warming shortens maturation and generation time of many microorganisms, and nutrients such as nitrogen and phosphorus stimulate growth. Along coastal areas worldwide, the discharge of sewage, industrial pollutants, eroding soil, and deposition of acid rain contribute effluents rich in nitrates and phosphorus which potentiate algal blooms (or eutrophication). Overharvesting of fish and shellfish combined with widespread pesticide use removes these consumers of plankton and contribute to overgrowth[42]. Additionally, sea level rise and altered storm patterns have the potential of enlarging bays and estuaries in which algae may proliferate[27]. In non-carbonated inland waterways, elevated atmospheric CO2 may directly stimulate the growth of algae and vascular aquatic plants. Increasing
UV-B radiation has been shown to harm some Antarctic algal populations[43]; however, adaptation to elevated UV-B radiation via pigment production has been observed[44]. Overall, during the past decade, phytoplanktologists report an unusual spread and intensity of algal blooms worldwide[45].
Algae, zooplankton, and vascular aquatic plants can serve as reservoirs for Vibrio cholerae and other enteric pathogens[46]. Large coastal blooms may have contributed to the multiple-epicentered pandemic in Latin America. Colwell and others[47] have demonstrated that quiescent forms of V. cholerae can persist on the sheaths and exoskeletons of marine organisms, and revert to an infective state when nutrients, pH and temperature permit. The recent isolation of chlorine-resistant, fully virulent, "rugose" or "wrinkled" colonial forms of V. cholerae in Peru, augments the potential for spread[48]. Toxic red tides, i.e., paralytic shell fish poisoning, diarrheic shell fish poisoning, and amnesiac shell fish poisoning also result from blooms of dinoflagellates. Ciguatera fish poisoning stems from the ascendance of a microalgal toxin through five trophic levels of the food chain[49].
Sea surface temperatures influence surface marine growth. One species of toxic phytoplankton, Gymnodinium breve, previously confined to the Gulf of Mexico, has resulted in dense red tides off the North Carolina coast[50]. This red tide, first reported in October, 1987 occurred after a "parcel of warm gulf stream water remained stranded on the North Carolina shelf," and resulted in human shellfish poisonings and substantial fishkills[50].
In the southern hemisphere in October, 1991, algal paralytic saxotoxins were detected in Southern Chilean waters and for the first time, as far South as the Straits of Magellan[51]. The unusual extension into new areas and its duration led to about 300 cases of intoxication from shell fish, including 11 deaths. This southern extension, coinciding with the penetration of cholera into Latin America, lends credence to the hypothesis that coastal algal blooms were associated with the intensity and breadth of the current pandemic. It now appears that environmental changes occurring on a global scale have already begun to alter the basic food web, and may compromise the safety of water and marine life consumption in temperate and tropical zones of the Americas.
STRATOSPHERIC OZONE DEPLETION AND PUBLIC HEALTH
The effects of chlorofluorocarbons (CFCs) on the stratospheric ozone layer were recognized in the 1970's, and the U.S. EPA banned these inert propellants in aerosols in 1978. By 1985, widespread concern erupted when an Antarctic-based British team discovered the "hole" in the ozone layer[52]. Subsequent passage of the Montreal Protocol in 1987, with amendments in 1990 and 1992, has already mandated sharp cuts in CFC production[53,54]. Yet, with half-lives of 70-110 years[6], CFCs' continued erosion of stratospheric ozone is anticipated through the next century. In April of 1992, NASA reported mid- to high-latitude stratospheric ozone levels 10-15% lower than any previous year, based on measurements by the "Total Ozone Mapping Spectrometer" (TOMS) on board the Nimbus-7 satellite[55]. NASA monitoring also detected high concentrations of CFCs and chlorine monoxide in the stratosphere above New England and eastern Canada[56]; it was only due to fortunate wind conditions dispersing these compounds, that an arctic ozone hole did not develop last spring.
Most ultraviolet radiation reaching the earth is in the UV-A range (320-400 nanometers). The atmosphere entirely blocks entry of the dangerous UV-C ( 180-290 nm) range of radiation. The stratospheric ozone layer absorbs much of the carcinogenic UV-B (290-320 nm) light[14]; UV-B radiation can induce cataracts, skin cancer, and immunosuppression, which may further influence cancer development and susceptibility to infection.
Cataracts
Ocular cataract formation causes half of the world's blindness and is associated with ultraviolet radiation in a "dose-response" relationship[57]. Amino acids and membrane transport systems in the lens of the eye are especially prone to photooxidation by oxygen radicals generated by UV-B irradiation[57]. Supporting this mechanism, dietary supplement with antioxidants such as -carotene and vitamin C, is reported from one study to lower the incidence of cataract extraction[58]. In an epidemiologic study involving Chesapeake Bay fishermen, direct personal exposure monitoring was implemented to control for varied protective measures (sunglasses and/or hats). The authors concluded that a doubling of UV-B exposure would cause a 60% increase in cortical cataracts over current levels[59].
Skin Cancer
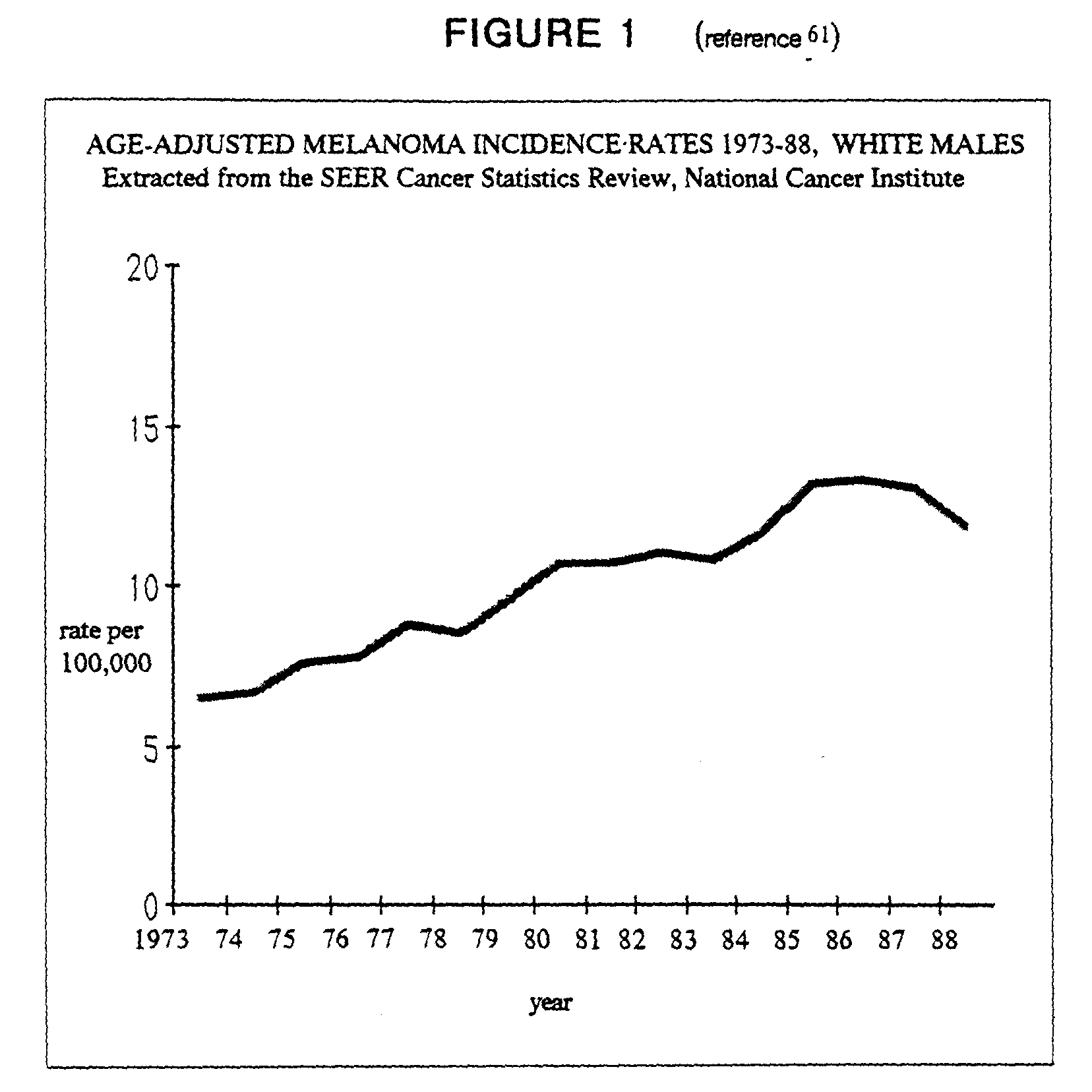
Nonmelanomatous skin cancer and superficial spreading melanoma are correlated with sunlight exposure[13,60]. In the U.S., malignant melanoma incidence among the white male population shows the steepest rise of all neoplasms, and from 1973 to 1988 increased by 86.3% (figure 1)[61]. In Scotland, Mackie, et al.,[62] report a rise of 82% (7.4% annually) from 1979 to 1989. While there is no direct relationship between cumulative ultraviolet exposure and melanoma, excessive UV exposure during childhood is associated with incidence[63]. For every 1% decline in the stratospheric ozone layer, a 3-5% rise in squamous cell carcinoma, 2-3% rise in basal cell carcinoma, and 1-2% increase in melanoma are expected[13].
Host susceptibility/ UVB-induced Immunosuppression
Immunocompromised individuals are at increased risk of infectious diseases and malignancies. Renal transplant patients on chronic immunosuppressive therapy, for example, have an increased frequency of skin cancer. Squamous cell carcinomas directly relate to sun exposure in these patients[64].
Pyrimidine dimer repair mechanisms play a central role in preventing carcinogenesis. Xeroderma pigmentosum (XP) is a rare, genetic condition involving a defect in this nucleotide excisional DNA repair system. A reduced capacity to excise UVB-induced pyrimidine dimers[65] leaves these patients at a 2000-fold risk of developing melanoma, and a 4800-fold chance of forming squamous and basal cell carcinomas by the age of 20[66].
A study by Ley, et al.,[63] supports the evidence for an inverse correlation between UV-B repair mechanisms and development of cutaneous melanoma. A "photoreactive repair" enzyme (activated by 320-500 nm wavelength light) in the South American opossum can split the pyrimidine dimers along a DNA strand, suppressing melanocyte hyperplasia. The authors conclude that UV-B can function as a complete carcinogen for the initiation and promotion of a malignant melanoma, specifically via DNA pyrimidine dimer formation[63]. (Inference from this photoreactive repair model might help explain why higher melanoma risk is related more to episodic compared to prolonged exposures)[67].
UV-B has been shown to inhibit cell-mediated immunity (delayed-type hypersensitivity) and immunosurveillance of transformed epidermal cells[64]. Several health consequences of this immunosuppression are recognized. HSV-I and II are reactivated by ultraviolet exposure, in a dose-response fashion[68]. UV-B can affect the course of infectious diseases of the skin such a onchocerciasis and dermatophytosis, and diseases where the skin is a portal of entry such as Leishmaniasis[14]. In sandfly-borne Leishmaniasis, parasite replication occurs in the dermis or epidermis, and is followed by hematologic dissemination and diffuse incurable disease[14]. In a murine model of cutaneous Leishmaniasis, Giannini found that low doses of UV-B irradiation increases the likelihood of systemic spread or reactivation of the protozoa[69]. Additionally, UV-B does not diminish the viability of parasites in the skin, and immunity to subsequent inoculations absent among animals irradiated with UV-B during the first infection[69]. These findings are consistent with diminished epidermal defense mechanisms and suggestive of reduced systemic immunity.
Suppression of tumor cell rejection has been specifically linked to UV-B, while assays using UV-A show no such suppression[70]. Moreover, in animal transplant studies, Daynes,et al.,[71] found that W-B-induced tumors fail to grow in normal recipient mice, but continue to progress when transplanted into irradiated mice.
Local and systemic effects
The ability of the immune system to function effectively depends on adequate antigen processing and presentation to T-cells, as well as augmentation of the response via lymphokine production. Langerhan cells contribute to delayed-type hypersensitivity skin reactions and may be responsible for processing of both microbial and tumor antigen[72,73]. Located above the basal layer of the epidermis, Langerhan cells gain no protection from melanin production by melanocytes[64], allowing low-dose UV-B to reduce their ability to effectively present antigens to T-cells[12].
Systemic effects are inferred by the observation that antigen-specific suppressor cells found in the spleens of UV-B irradiated mice[74] are stimulated in a dose response relationship[75]. In vivo, W-B can impair natural killer cell activity against a melanoma cell line[76] at doses comparable to those found in the environment[77]. These findings are consistent with the epidemiologic associations between ultraviolet exposure and melanoma.
In addition, UV-B stimulates keratinocytes to secrete interleukin- I (Il-1), which in turn, stimulates pituitary production of adrenocorticotropic hormone (ACTH), and subsequently, glucocorticosteroid levels. Reduced Il-2 and elevated Il-4 have also been observed, paralleling the profile one sees following steroid administration, and further substantiating the presence of UV-B induced alterations in the cascade of immune responses[12].
UV-B impact on vaccine take and efficacy is largely unexplored. A study in mice involving BCG has shown that delayed-type hypersensitivity is retarded initially, but returns under continued UV-B exposure[78]. This suggests that adaptive measures, such as increased skin thickness, could dampen UV-B's interaction with humoral defense.
Sunscreens and UV radiation
Erythema is a measurable biological endpoint of UV exposure and is used to determine sunscreen effectiveness[79]. "Sun Protection Factor"(SPF) is rated by the length of time required to elicit a "minimal erythematous dose" (MED)[80]. Sunscreens have also been shown to partially block the immunosuppressive response, however, in no correlation with their SPF designations. While histologic signs of erythema are blocked by sunscreen application, the protective agents may fail to prevent UV-B induced tumor susceptibility[80]. One study[81], suggests that the dose of UV-B necessary to increase animal susceptibility to cancer by immunosuppression is 10-fold less than the amount needed for overt carcinogenesis by molecular damage.
Exposure to UV-B in tanning salons can cause subsequent failure to respond to antigen challenge[82], and suppress enzyme repair of DNA lesions. A major public health concern is that by reducing the erythematous reaction, sunscreens may deceive people into remaining exposed to solar rays and their immunosuppressive effect for longer periods.
Measuring DNA nucleotide excisional repair may be capable of predicting subpopulations at risk of developing skin cancer. An assay has been developed to measure this mechanism, which is found to be significantly reduced in skin cancer cases[83]. Xeroderma pigmentosum patients have provided a model for testing this assay, for the defect in their repair mechanism phenotypically varies considerably within this subpopulation.
Ambient Ultraviolet Levels
A dose of UV-B as small as 0.15 - 0.30 kilojoules/m[2] can damage epidermal Langerhan cells[14]. 22-34 kJ/m[2] can alter lymphokine profile[49], kJ/m[2] inflicts systemic suppression, and 86 kJ/m[2] leaves the immune system unresponsive to UV-induced tumors[14]. At latitudes between +25deg. N and -25deg. S, ambient midday UV-B flux on a clear day could conceivably expose a person to up to 86 kJ/m[2] during a 10 hour period[14].
One can expect rising ambient UV-B levels from stratospheric ozone depletion. Robertson-Berger meters at U.S. National Weather Service stations have been recording UV-B levels since 1974[84]. Readings are weighted according to the action spectrum producing erythema, the most bioactive wavelength being 297 nm. Ambient levels vary depending on latitude, altitude, cloud cover, and air clarity. From 1974 to 1985, no positive trends in surface UV-B were found[84]. However, only 25 recording stations existed in the U.S. as late as 1990, and were located primarily at large airports[85]. Anthropogenic tropospheric ozone and other aerosols released in urban areas have likely impeded accurate UV-B measurement[86]. A high altitude station in Switzerland (far removed from the influence of air pollution) has observed increases in ambient UV-B levels[87].
RECOMMENDATIONS
Considering the current and potential health consequences of global climate change, primary prevention through environmental protection is essential. The health outcomes in temperate regions presented in this paper represent only a partial evaluation of the predicted effects of global climate change. Repercussions such as sustained crop failures or coastal flooding could be especially catastrophic for countries with inferior infrastructures.
World consensus, as expressed at the UNCED meetings in June, 1992, emphasized the need to promote sustainable development while conserving natural resources. Our recommendations towards this goal include: 1) reducing greenhouse gas emissions by providing incentives for better energy efficiency and renewable technologies; 2) preserving tropical rainforests through improved agricultural policies (shifting away from meat production), economic diversification, and vastly expanded "debt-for-nature" swaps; 3) replacing CFCs with alternative refrigerants and transferring such technology to developing countries; and 4) slowing population growth through comprehensive plans, including rural development and universal education for women.
Internationally, initiatives such as the Montreal Protocol have already established strict guidelines for cuts in CFC production. UNCED helped to advance cooperation among countries, non-governmental organizations, and the United Nations in establishing guidelines to reduce other anthropogenic causes of rapid climate change and instability; incentives for equitable and environmentally sound growth are the main goals of future collaboration. International groups such as the World Meteorological Organization and the International Council of Scientific Unions are also intensifying activities to study global climate change[86]. On the national scene, Vice President, Al Gore, has proposed to reduce greenhouse gas emission by levying a tax on major carbon producing industries[88].
Because climate changes may, however, already be out-pacing our response, secondary prevention through early detection is also necessary to avert or minimize deleterious health effects. Monitoring of skin cancer, infectious disease, and other parameters requires multidisciplinary support and coordination. Interpreting climatological data bases to predict regionally specific public health outcomes will be difficult and must incorporate cyclical weather patterns, sunspot activity, volcanic eruptions, and uncertainties in modeling. For example, El Chichon and Mount Pinatubo produced enough particulates and sulfur dioxide to temporarily lower global temperatures back to levels of the 1960's[89]. Climate models contain uncertainties concerning "sinks" for CO2, ocean heat capacity, reflectivity of polar ice caps, and the net and regional impact of clouds[6].
Improving exposure assessment will be key to any endeavor attempting to detect diseases of environmental etiology. To address increases in ambient UV-B, meters should be placed in rural locations, thereby lessening the influence of local air pollution (particularly at airports) and at locations of high risk behavior, such as beaches. Robertson-Berger meters, weighted only for 297 nm, may fail to detect other significant UVB wavelengths[44]. Meaningful exposure assessment will be best accomplished by selective use of personal UV monitoring devices. Immediate preventive measures should parallel new monitoring efforts, including ultraviolet advisory warnings (already operational in Australia) and an Occupational Safety and Health Act (OSHA) standard for UV-B exposure to outdoor laborers[86].
Epidemiologic studies to evaluate key indicators such as age-adjusted cataract rates or squamous cell carcinomas in a select sentinel population such as outdoor laborers, should be conducted in regions where elevated ambient UV-B exposures have already been documented. Incidence of Herpes simplex and zoster reactivation may also provide markers for immunosuppression. Laboratory research into the human immune system's functional response to UV-B, i.e., vaccine efficacy, is needed, as are animal studies identifying wavelength-specific mutations associated with malignant changes.
Surveillance provides valuable data only in conjunction with comprehensive reporting[86]. Tumor registries must be expanded to accept all non-invasive, nonmelanomatous skin cancers. The U.S. has 38 state cancer registries, but no centralized national linkage. The U.S. National Cancer Institute has initiated coordination by creating the Surveillance Epidemiology and End Results (SEER) registry in 11 of the 38 registries[90]. Cancer diagnoses are based on pathologists' findings[91], therefore, sentinel laboratory-based active surveillance of UV-B induced skin cancer at the 11 SEER sites would serve as a rational step to standardize monitoring.
To track vector-borne diseases, four types of surveillance are possible: 1) tallying human cases; 2) determining distribution and infectivity of vectors; 3) monitoring a broad range of non-human vertebrate reservoir species; and 4) following weather patterns to help predict vector distribution[41].Testing of one reservoir species for SLE is already in place in Florida, where seroconversion rates in "sentinel chickens" are monitored in each county[92]. Excessive winter rainfall combined with summer droughts have commonly preceded SLE epidemics, and so makes weather monitoring useful as well. For diseases related to coastal contamination, international monitoring is being developed by the International Oceanographic Commission of UNESCO. Further efforts in the area of infectious disease should include: 1) educational campaigns sensitizing physicians and state health departments to achieve more thorough ascertainment of related cases; 2) more laboratory and field investigations of arthropod vector ecology and pathogen infectivity at elevated temperatures; and 3) coordination of infectious disease reporting between private and public laboratories to more rapidly detect the emergence of new diseases and/or the redistribution of old ones.
Computerized data bases and Geographic Information Systems (GIS) can greatly assist in this process. A diskette database developed by The Institute for Scientific Information entitled, "Focus on Global Change", surveys reports in 8000 journals[93]. The United Nations' "Infoterra", for example, compiles environmental information affecting 99% of the world's population[94]. An expanded global health monitoring system, coordinated through the World Health Organization, is in the developmental stages.
This medical review has focused on the public health outcomes of global climate change for temperate regions. The chief aim for evaluating these consequences is to promote primary prevention. Yet, considering political and economic inertia, efforts to abate greenhouse gas emission and CFCs through national and international policies may not progress rapidly enough to eliminate the environmental risks already set into motion. The medical and public health profession must participate cooperatively with environmental scientists and regulators to assess and confront the complex threat of accelerated climate change. The healthcare community will be called upon further to educate the public and lawmakers concerning the "costs" of inaction and the "benefits" of bold measures to achieve primary prevention. Public health must be central to the policy making process, as health is the true measure of national and international security.
ACKNOWLEDGEMENTS:
We gratefully acknowledge Robert E. Shope, MD, for his critical review of the manuscript. We also extend thanks to Bernard Goldstein, MD, Lee Harrison, MD, and Jean Brennan, MS for their helpful recommendations.
REFERENCES
5 Epstein PR. UNCED: a conference report. PSR Quarterly 1992 Sept; 2(3): 177-81.
25 Spielman A, (personal communication).
32 Halstead SB. Pathogenesis of dengue: challenge to molecular biologists. Science 1988; 239: 476.
37 Shope RE. Arbovirus-related encephalitis. Yale J Biol Med 1980; 53:93-99.
50 Tester P. Red tide: effects on health and economics. Health and Environ Digest 1991: 5: 4-5.
51 Lembeye G. Major PSP outbreak in Chile, 1991- 1992. Harmful Algae News 1992; 2: 1-2.
56 Kerr RA. Ozone hole not over the arctic - for now. Science 1992; 256: 734.
72 Wolff K, Stingl G. The Langerhan cell. J Invest Dermatol 1983; 80(6) Suppl:17 S-21 S.
88 Gore A. Earth in the Balance. Boston, Houghton Mifflin, 1992.
89 Kerr RA. Pinatubo global cooling on target. Science 1993; 259- 594.
91 Muir CS. The cancer registry in cancer control: an overview. IARC Scientific l985; 66: 13-26.
93 Institute for Scientific Information. Letter. Philadelphia, PA, 1992.
{kind=link}
{kind=link}