 Reproduced, with permission, from:
Liehne, P. F. S. 1988. Climatic influences on mosquito-borne diseases in Australia. In Greenhouse: Planning for climate change, ed. G.I. Pearman. Australia: CSIRO.
Reproduced, with permission, from:
Liehne, P. F. S. 1988. Climatic influences on mosquito-borne diseases in Australia. In Greenhouse: Planning for climate change, ed. G.I. Pearman. Australia: CSIRO.
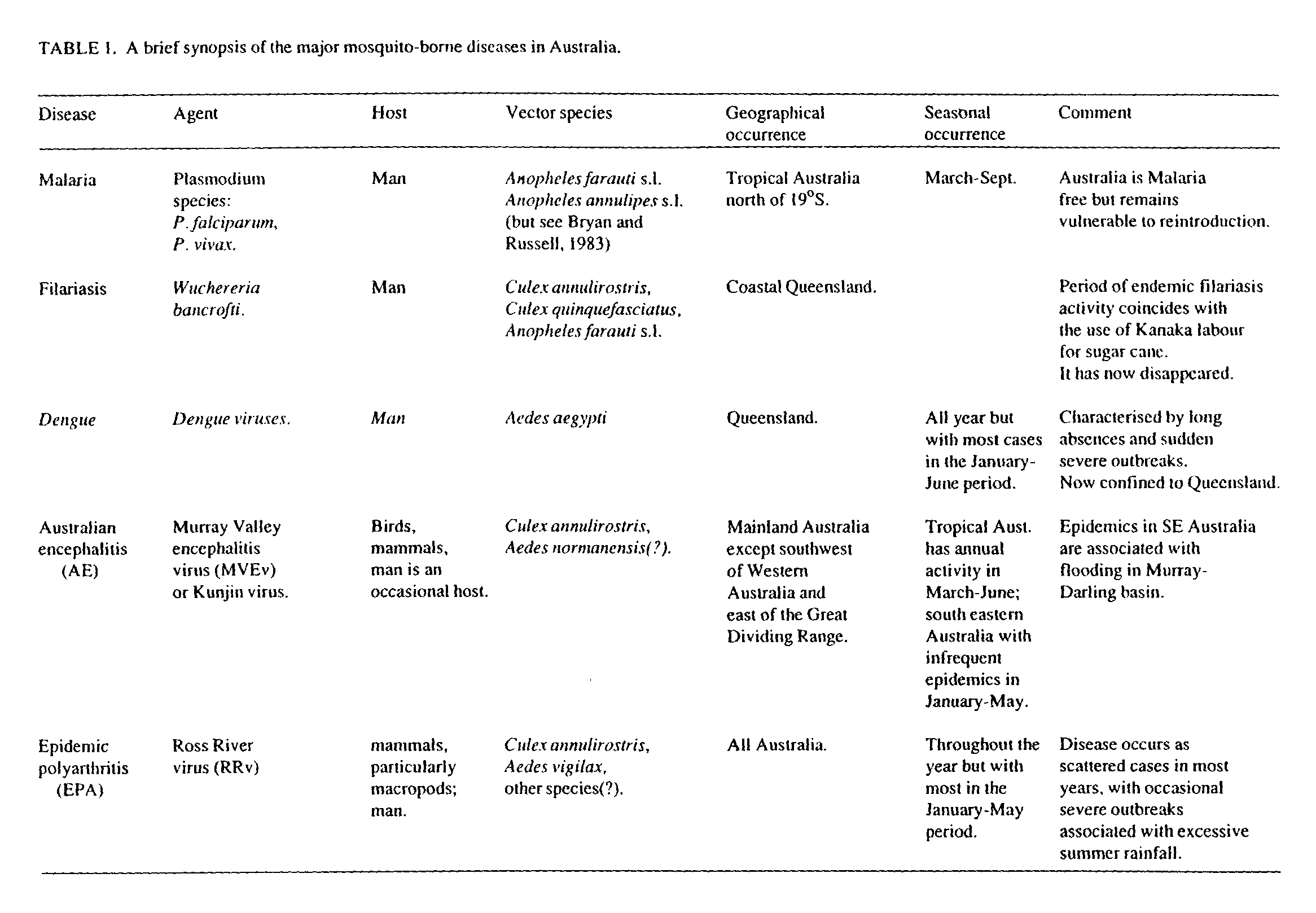
The most important mosquito-borne diseases in Australia are the human arbovirus dengue, and the zoonotic arboviruses Murray Valley encephalitis virus (causing Australian encephalitis) and Ross River virus (causing epidemic polyarthritis). Historically, the parasitic diseases malaria and filariasis have also been significant.
If the greenhouse effect increases rainfall and temperature, it can be expected to influence the seasonal and geographical abundance of the major vector species and vertebrate hosts. In tropical Australia, the impact of these changes should (a) increase receptivity and thence vulnerability to malaria, (b) increase the incidence of epidemic polyarthritis, and (c) extend the geographical area of endemicity of Murray Valley encephalitis virus and possibly the frequency of Australian encephalitis outbreaks, particularly at the southern boundary of the monsoonal influence. It may have an effect on the incidence of dengue, but this could be limited as the vector mosquito breeds in domestic container habitats which are often artificially maintained by man, and are thus independent of rainfall.
In temperate Australia, the increased winter temperatures and summer rainfall will extend the season favourable to Culex annulirostris, the dominant vector species. Breeding of this species could extend from mid-September to mid-May permitting much greater summer populations to be reached. This could favour increased incidence of epidemic polyarthritis. Possible changes to the incidence of Australian encephalitis are less clear. If increased spring and summer rainfall permit the required threshold associations of vectors and hosts to be exceeded more frequently, Australian encephalitis outbreaks/epidemics may become more frequent. On the other hand, if this threshold is exceeded only in those years with major springfloods associated with intense El Nino/Southern Oscillation variations, then the incidence of Australian encephalitis epidemics will depend on the changes to the El Nino/Southern Oscillation resulting from the greenhouse effect.
Key word index: arbovirus, Australia, climate, temperature, rainfall, greenhouse effect, ENSO, Murray Valley encephalitis virus, Ross River virus, dengue, malaria, Australian encephalitis, epidemic polyarthritis.
Mosquitoes cause significant nuisance throughout much of Australia and are responsible for transmission of a number of human diseases. The arboviruses are perhaps the most important diseases in the Australian context. These include Australian encephalitis (AE, formerly known as Murray Valley encephalitis), epidemic polyarthritis (EPA, often misleadingly referred to as Ross River fever) and dengue. Two parasitic diseases (malaria and filariasis) have been historically important and malaria has the potential to re-establish should it be re-introduced into susceptible areas.
A number of studies have recognised the relationship between arboviral disease and climate (Davies et al., 1985; Foo et al., 1985; Grady et al., 1987; Mogi, 1983; Monath, 1980). In Australia, the incidence of epidemic AE has been linked to climatic factors, particularly rainfall and flooding (Anderson and Eagle, 1953; Forbes, 1978; Miles and Howes, 1953).
In this paper, I shall review the climatic influences on the dynamics of vector populations and vector-borne disease transmission. The discussion is, of necessity, simplified, but provides a framework within which to discuss the impacts of the greenhouse effect. Whilst the discussion is limited to mosquito vectors and the diseases which they transmit, the conclusions may be extrapolated to pest species which have similar larval requirements. It should be noted that there is great local variability in respect of climatic factors, topography and physiography, and that mosquito diversity and seasonal abundance. and thence disease transmission, can vary accordingly.
Mosquito-borne diseases require that a mosquito pass the pathogen from one vertebrate host to another, and successful transmission requires appropriate aggregations of vectors and vertebrate hosts. Where the disease is strictly human, the close association of vector and man is essential. For zoonoses, the natural cycle involves the vector and a vertebrate host, and man may become infected when he accidentally enters this transmission cycle, or when these cycles become established near to human habitation.
All the diseases discussed in this paper have an obligatory cycle of replication and/or development within the mosquito vector. The extrinsic incubation period (EIP) defines that period of development within the vector between first ingestion of the disease organism and when it can be transmitted. The EIP is temperature dependent and varies between mosquito vector species and for different viruses or parasite species. For example, 50-90% of Culex annulirostris can transmit Murray Valley encephalitis virus (MVEv--one of the viruses causing AE) after 10 days extrinsic incubation at 28deg.C (Kay et al., 1984b). A general extrinsic incubation period for malaria is 9-15 days (Service, 1980). It is generally accepted that once the EIP is passed, the vector remains infective and can transmit the disease for the remainder of its life. It is the length of the EIP that makes factors such as vector longevity and population density so crucial to successful transmission. In simple terms, the greater the number of vectors and the longer they survive, the greater the chance that an infected vector will survive to become infective, feed on a susceptible host and so transmit the disease.
The vector status of mosquito species which are biologically competent to transmit the disease agent is determined by a number of factors. For example, fecundity, feeding preference and specificity, oviposition site selection (ie. larval habitat preference) and dispersal can determine the potential for vector population growth, the degree of contact of vector and host, and the capacity for spread of the disease.
For zoonoses, the degree of vector/host contact, the level of herd immunity in the vertebrate host, recruitment and survival in the host population and the availability of suitable alternative hosts can also be critical to successful transmission.
Climatic factors, particularly temperature and rainfall, can have a profound impact on transmission cycles by influencing the availability of vector breeding sites, extending vector longevity, altering host breeding or migration patterns, and maintaining aggregations of vectors and hosts around water bodies.
Table 1 presents a brief analysis of the mosquito-borne diseases of human significance in Australia, and Table 2 presents a brief analysis of the biology of the major vector species. Malaria and filariasis are obligatory parasites of man. Dengue, a flavivirus, is also a disease of man. Fortunately, the major vector species for these diseases are largely restricted to sparsely populated areas in tropical Australia. The recent outbreak of dengue in Queensland (1982-85) (Kay et al., 1984a) clearly indicates that transmission can occur following re-introduction of the disease into localities with a susceptible human population and where the vector is present. These tropical diseases are widely distributed in the neighboring South East Asian and Western Pacific Regions. AE and EPA are zoonotic diseases indigenous to Australia and New Guinea.
Malaria
Malaria is a parasitic disease of man caused by infection with members of the genus Plasmodium. Australia is considered to be malaria free, but tropical areas remain vulnerable to the re-introduction of malaria. The disease is transmitted by mosquito species belonging to the genus Anopheles, particularly Anopheles farauti s.l..
An farauti s.l. is recognised to be a sibling species complex with at least three members (identified as An farauti No. 1, No.2 and No.3) reported from Australia (Mahon and Miethke, 1982). The distributions of the three member species in Australia is unclear. An farauti No. 1 is recognised as the major malaria vector in Papua New Guinea and presumably also within Australia, but little is known of the vector status of the other two members of the complex (Bryan and Russell, 1983). Bryan and Russell review the vector status of other Australian anophelines and conclude that Anopheles bancroftii is a secondary vector, whilst other species are considered incidental or suspected vectors. An farauti No. 1 breeds in brackish coastal swamplands along the tropical coastline from Townsville to at least Darwin, but may extend as far as Wyndham. An bancroftii is similarly a dominant species in the semi-arid tropics, breeding in semi-permanent to permanent, vegetated fresh water habitats. Both species are most abundant in the early to mid dry seasons, when the respective breeding sites are most prevalent. For An farauti No. 1, this coincides with the period when ground water and run-off dilutes and maintains tidal wetlands as semi-permanent brackish swamps. For An bancroftii this period is when the vegetation growth in fresh water swamplands maintained by wet season rainfall run-off and raised water-tables has become well established and provides shelter and protection to the larvae.
Other anophelines, whilst not considered to be major vectors, may have the capacity to transmit malaria under conditions of poor housing and hygiene.
Filariasis
Filariasis is an historically important parasitic disease in Australia. It was particularly prevalent in the sugar-growing areas of Queensland in the early part of this century. Its origins and eventual disappearance can be linked to the arrival and subsequent repatriation of the Kanaka labour used to develop the cane industry.
In Australia, the causative filarial worm was Wuchereria bancrofti, and the vector was Culex quinquefasciatus (= fatigans), the common urban pest mosquito. Cx quinquefasciatus breeds in fresh to polluted domestic waters and is widely distributed throughout Australia.
Filariasis has not been recorded endemically from Australia for many years, and it is unlikely that it will become re-established even if re-introduced.
Dengue
Dengue is caused by infection with one of the four flaviviruses (dengue 1 - dengue 4) which make up the dengue group. In Australia, man is the only known vertebrate host, and the vector is the domestic container breeder, Aedes aegypti. Dengue occurs as infrequent, but occasionally severe outbreaks/epidemics. Recent epidemics have been restricted to Queensland, occurring in 1953-55 and 1981-82 (Kay et al., 1984a), and the disease was wide spread in tropical Australia during the second world war. Only the non-lethal form of the disease (dengue fever) has been reported in Australia this century, but the severe forms (dengue haemorrhagic fever and dengue shock syndrome) are of great concern in Asia, the Philippines and Indonesia, and probably occurred in Queensland late last century. It appears that dengue is re-introduced into the receptive areas of Australia via infected people returning from the endemic areas of Asia or the Western Pacific.
Ae aegypti, though formerly widespread throughout tropical Australia, is currently restricted to Queensland, particularly in north-east coastal areas. It breeds in a variety of domestic container habitats, and recent surveys indicate that plant containers and garden receptacles form the most significant breeding sites (Kay et al., 1987). This is particularly important as these breeding sites are often maintained by human activity and are largely independent of precipitation.
Australian encephalitis (AE)
AE is an encephalitic disease caused by infection with the flavivirus MVEv. There is a high rate of asymptomatic infection, but the outcome of symptomatic infection can be severe. Approximately one third of AE cases recover fully, one third are expected to die from the disease and one third suffer residual sequelae which vary from minor personality and psychological disorders, to severe paralysis and brain damage. It has recently been shown that the related flavivirus, Kunjin virus, can cause a syndrome indistinguishable from infection with MVEv. Kunjin has essentially similar ecology to MVEv, and the following discussion will be restricted to the more important virus.
MVEv survives in zoonotic transmission cycles involving a range of vertebrate reservoir hosts, particularly birds. The dominant vector is Cx annulirostris. In discussing zoonoses such as AE, it is necessary to differentiate between the disease and the causative virus. It is also necessary to evaluate the differing patterns of activity in tropical and temperate Australia.
Tropical Australia
MVEv is enzootic in tropical Australia with annual activity recorded in the Kimberley region of Western Australia, generally in the March/June period (Liehne et al., 1976; 1981; Wright et al., 1981). Activity of MVEv is recorded less frequently (about every 3 to 10 years) from the Pilbara region of Western Australia, the Northern Territory and Queensland. The dominant vector is Cx annulirostris, though the recent isolation of several strains of MVEv from field caught Aedes normanensis has indicated that this species may be a vector in the Kimberley region of Western Australia, and may be important in the survival of the virus through adverse seasons.
Cx annulirostris breeds in the vegetated margins of semi-permanent and permanent fresh wetlands, and also in organically polluted sites. It breeds throughout the year, but with peak populations in the period following the wet season (March to June) when the breeding sites are most extensive and stable. This coincides with the period when most arboviruses are isolated from field caught mosquitoes.
In those tropical areas where monsoonal rainfall is reliable, Cx annulirostris is the dominant species of the mosquito fauna. Further south, where the monsoonal rainfall occurs as scattered thunderstorms, the species survives in residual populations breeding in the few permanent water bodies which form a refuge. The species, however, will colonise any semi-permanent sites created by local rainfall and the populations may be very high on occasion.
Ae normanensis, on the other hand, is a floodwater mosquito found in the semi-arid tropics. The species has drought resistant eggs which remain dormant within the soil substrate of the temporary, rain-filled depressions which form the main breeding sites. The eggs hatch when the breeding site is filled and populations generally rise rapidly following local rainfall. Resurgence of the species can be noted if there is some delay between successive rain storms. The species is predominant in regions where the rainfall is erratic, that is, in the drier portions of the monsoon zone where rain occurs as scattered thunderstorms and in areas which receive the majority of rain from infrequent cyclonic activity.
AE occurs as frequent small outbreaks (some consisting of only one or two cases) in tropical Australia (Anon., 1987). Cases are recorded from widely scattered localities in the March to June period (see Bucens, 1982; Stanley, 1982) every 2 to 5 years. The frequency of AE in the tropics reflects the activity patterns of MVEv. Cases occur more frequently in the Kimberley region. and are less frequent in the Pilbara region, the Northern Territory, and Queensland. The disease is generally less severe in the tropics than in epidemics in temperate south east Australia.
Temperate Australia
It is considered that MVEv may also be enzootic in south east Australia in cryptic, as yet undefined. cycles. Overt transmission has only been recorded in years of excessive rainfall and severe flooding in the Murray-Darlino basin (Forbes, 1978), and is generally indicated by human cases of AE. Numerous strains of MVEv were isolated from field caught mosquitoes during the epidemic of 1974 (Marshall et al., 1982b), but no strains of this virus have been recovered in subsequent years (Marshall, pers.comm.). This is in stark contrast to the situation in the Kimberley region of Western Australia where the virus can be isolated from mosquitoes in each year (Liehne et al., 1976; 1981).Neither MVEv nor AE has been recorded from the south west of Australia.
Infrequent, but severe epidemics of AE are recorded from south east Australia (Forbes, 1978; Stanley, 1982). The first case in the two most recent epidemic years (1951 and 1974) occurred in the first week of January and the epidemics extended into April/May (Anderson, 1952; Forbes, 1978). Whilst these epidemics were centered on the Murray Valley, cases were recorded from more northerly sites as the epidemics progressed, particularly in 1974 (Marshall, 1979). Smaller outbreaks in south east Australia were recorded in 1956 and 1971. yielding 3 and 1 case respectively (Forbes, 1978; Stanley, 1982; Anon., 1987).
Following the 1951 epidemic of AE, the Miles/Anderson theory was proposed to explain the sporadic nature of the epidemics. MVEv was considered enzootic in northern Australia (northern New South Wales? to New Guinea), and the extensive flooding seen in epidemic years permitted the movement of the virus, probably via infected birds, into the Murray Valley (Miles and Howes, 1953; Anderson and Eagle, 1953). The north-south movement of MVEv has been questioned by Marshall (1979), who notes that the recorded cases in epidemics indicate a south-north progression.
Sellers (1980) reviewed the occurrence of a range of arboviruses. including MVEv, and argues that MVEv is enzootic in tropical zones (probably in New Guinea) and is re-introduced into epidemic zone via wind-borne infected vectors or vertebrate hosts when conditions are suitable. Sellers offered no evidence to support his argument, and did not review the detailed data available from either the 1951 or 1974 epidemics. However, detailed analyses of the spread of bovine ephemeral fever, a veterinary arbovirus transmitted by Culicoides species and possibly also Cx annulirostris, has clearly demonstrated that the southward movement of the virus was closely correlated to prevailing weather patterns (Murray, 1970; Newton and Wheatley, 1970).
Forbes (1978) analyzed rainfall patterns and the incidence of AE. He concluded that transmission of MVEv and the occurrence of AE cases in south east Australia was in the second of two successive years with summer rainfall in excess of decile seven in all five main eastern watersheds. Grady et al. (1978) defined similar predisposing rainfall conditions over the preceding summer for the occurrence of Eastern equine encephalitis in Massachusetts.
Rainfall in south east Australia is correlated to Variations in the El Nino/Southern Oscillation (ENSO) and Nicholls (1986) has developed a predictor of AE activity based on the mean Darwin barometric pressure.
The major vector in south east Australia is Cx annulirostris. However, in temperate Australia, the seasonal activity of this species is restricted to the warmer summer months. The species overwinters as quiescent adult females (Russell, 1987) which become active again only when the mean ambient temperature reaches 17.5deg.C (McDonald, 1980; Russell, 1986a), generally from November to March. The species breeds in the vegetated margins of fresh water habitats. In the Murray Valley, the most important breeding sites are largely maintained by the management of waters for flood mitigation and irrigation.
The impact of rainfall and local flooding on mosquito breeding can vary according to local geography, but generally higher populations are recorded with more extensive flooding. Russell (1986b) discusses these influences on mosquito populations at Echuca between 1979 and 1985 and shows that the highest populations recorded were in the summer of 1983/84, coinciding with the greatest degree of local flooding. However, overall mosquito densities at Echuca remained well below that of the epidemic year 1973/74. The major contributing factor appears to have been the persistence of spring flood waters through to February in 1974 (Russell, 1986a).
As in the tropics, there appears to be a general correlation between population densities of mosquito vectors and the recorded activity of arboviruses. The summer of 1973/74 recorded extremely high densities of mosquitoes (Marshall et al., 1982a) accompanied by widespread and intense activity of arboviruses (Marshall et al., 1982b) and 58 cases of AE were reported (Forbes, 1978). Similarly, the less extensive floods of the 1983/84 summer resulted in the highest mosquito populations between 1979 and 1985, accompanied by relative increase in arbovirus activity (Russell, 1986a,b). and coincided with a major outbreak of EPA in New South Wales (Hawkes et al., 1985).
In most localities, mosquito populations in the summer of 1983/84 were of sufficient densities to support intense and widespread activity of arboviruses (Marshall, pers.comm.). That these high populations of vectors were not accompanied by transmission of MVEv and the occurrence of AE requires some explanation. The answer may lie in the activity of vertebrate hosts. The summer of 1973/74 triggered an extraordinary explosion of wildlife throughout Australia, and particularly of waterbirds (Marshall et al., 1982c). Such overt and extensive waterbird breeding was not recorded in 1983/84. One may speculate that above average summer rainfall in the year preceding epidemic activity of AE is required to prime the system. Wildlife, particularly waterbirds, may respond to the second year of optimum conditions by intense breeding. The long lead times of barometric indicators (see Nicholls, 1986) may also trigger wildlife to respond by extensive breeding and migratory activity.
The question whether MVEv is reintroduced into the Murray Valley during the epidemic summer or in the previous summer, or whether it survives there in cryptic cycles remains unresolved.
Epidemic polyarthritis (EPA)
EPA is characterised by arthritis, rash, and constitutional symptoms such as fever, fatigue and myalgia. It is caused by infection with the alphavirus Ross River virus (RRv). RRv is enzoonc throughout Australia and activity is generally greatest in the summer and autumn. In most years, EPA is recorded as geographically scattered cases throughout the year, but with the preponderance of cases in the January to May period, particularly in the tropics. Occasional severe outbreaks have been recorded in temperate Australia following extensive summer rainfall (Hawkes et al., 1985).
The accepted major vectors of RRv are Cx annulirostris and Aedes vigilax. The virus has been isolated from a range of mosquito species from several genera, and these may also be vectors in some circumstances. The natural hosts for RRv are marsupials. particularly macropods. Man has also been shown to be an effective host in epidemic situations in the South Pacific.
The ecology of Cx annulirostris has been discussed in some detail above. Ae vigilax is a very common coastal pest species. It has a drought resistant egg and breeds in tidal salt marshes. The highest populations are usually recorded during November to January. This period coincides with the period of peak spring tides, and the absence of rainfall. In temperate areas, the tidal wetlands are maintained as brackish swamps through the winter and are saline following inundation by spring tides in the November/January period. In the tropics, the onset of the monsoon in December/January marks a decline in populations of Ae vigilax as rainfall flushes out tidal pools and maintains these wetlands as brackish swamps more suitable to species such as An farauti No. 1.
It is widely believed that the increasing atmospheric concentrations of greenhouse gasses over the next half century will result in significant climatic changes. Many general observations can be made, for example regarding ocean warming, sea-level rises and changing atmospheric temperatures. However, the resolution of climate models is such that regional scenarios are generally inaccurate. This results from the inability of available models to integrate and define all relevant climatic processes, and the limitations of current computer technology which can only allow resolution of physical processes over two orders of magnitude (Schlesinger and Mitchell, 1987).
A recent comparison of the predictions from three general circulation models to a doubling of atmospheric carbon dioxide gave quite contradictory results for the Australian region (Schlesinger and Mitchell, 1987). For example, summer rainfall predictions varied from a decrease of up to 2 mm d[-1] over much of eastern Australia; to little change over northern Australia and a slight decrease in the south east; to slight increases over much of Australia with the greatest increases (up to 2 mm extra per day) being in the north west of Queensland. Similar anomalies were seen for winter rainfall, soil moisture and winter surface air temperature predictions.
Pittock and Salinger (1982) and Pittock (1983) have developed a relatively detailed projection of climatic conditions expected in Australia following a doubling of atmospheric carbon dioxide. Despite these authors' caveats, and the obvious contradictions in predictions from different general circulation models, this scenario provides a framework for evaluating the impact of the greenhouse effect on vectors and vector-borne disease in Australia.
The main features of the scenario developed are (a) increased temperatures of 2deg. to 4deg. with maximum warming in the south and in winter; (b) higher spring, summer and autumn rainfall over much of eastern Australia and particularly the Murray-Darling basin; (c) more intense and greater penetration of monsoonal rains in northern Australia and a lengthening of the monsoon by two to four weeks; (d) a general rise in sea level of 0.2 to 1.4 m; and (e) a greater frequency and intensity of tropical cyclones, and a shift southward of some 200-400 km for this cyclonic activity (Pittock and Salinger, 1982; Pittock, 1983; Appendix, this volume).
In tropical Australia, the impact of the proposed scenario will favour the following changes. The geographical area where the mosquito populations are dominated by Cx annulirostris and Anopheles species will extend southward as previously temporary wetlands become more permanent. The southward extension of monsoon associated thunderstorm activity would result in an extension of the pest problems resulting from floodwater Aedes such as Ae normanensis.
Near the coast, sea-level rises will enhance the penetration of tidal saline waters into estuarine and tidal river habitats, probably extending into the flood plains of these rivers. This would exacerbate breeding of Ae vigilax and An farauti No 1 close to urban areas, and expose greater numbers of people to these pest and vector species. The magnitude of such changes is difficult to determine as, if the changes occur gradually, new drainage patterns may establish to reduce breeding potential in the newly created sites.
A complicating factor may be the problem of cyclonic storm surges. The frequency of major storm surges will increase leading to greater inundation of low lying areas and a further exacerbation of pest and vector mosquito breeding in these sites.
The impacts of these changes will provide a generally southward extension of currently observed arbovirus activity. Prolonging the rainfall season could delay the onset of arbovirus activity but may extend transmission later into the year. Accordingly, MVEv transmission may be enhanced by the greater populations of the vectors, and this may exacerbate MVEv activity in the Pilbara and Gascoyne regions of Western Australia. There may be a corresponding increase in the incidence of AE. Similar effects may be observed in Queensland and the Northern Territory.
The increased populations of both Cx annulirostris and Ae vigilax will enhance the potential for transmission of RRv and may increase the incidence of EPA.
The problem of dengue in Queensland may not change significantly as the vector (Ae aegypti) breeds in garden and household receptacles which are often maintained by human activity. Whilst exacerbation of Ae aegypti breeding in rain-filled containers would be expected, the artificially maintained populations should be sufficient to support transmission of dengue if it is re-introduced. That is, there may only be a slight increase in the potential for dengue transmission.
The potential for re-establishment of malaria will be increased as the vector populations adjacent to human habitation are enhanced, and the geographical limits of the malaria receptive zone may be extended to the south following expanded activity of the vectors.
In temperate Australia, the higher winter temperatures would allow an earlier commencement of mosquito activity. The 17.5deg.C threshold for Cx annulirostris breeding will extend from mid-September to mid-May, potentially permitting the development of very much larger spring and summer populations. The higher rainfall in spring and summer, particularly in the Darling River basin, will result in more frequent local flooding, and may permit more frequent and intense breeding of wildlife, particularly birds. The degree to which this occurs will depend on the capacity of the system of river, flood and irrigation management to deal with the altered pattern of river flows. The reduced levels of snow pack will decrease the volume of run-off in spring and may reduce the threat of flooding in the Murray River.
Near the coast, increased sea levels may allow populations of coastal breeding mosquitoes, particularly Ae vigilax, to increase as tidal penetration of low lying areas is extended.
The impacts of such changes on disease transmission would be that the background levels of transmission in 'normal' years would be enhanced. That is, the potential for transmission of RRv will probably be greater in both geographical and seasonal extent, and incidence of EPA infections may also rise.
The impact on the prevalence of AE is less clear. AE currently occurs in years of major spring/summer flooding when the critical thresholds of vectors and hosts are exceeded. If the impact of the greenhouse effect allows the critical thresholds to be exceeded more frequently, then the activity of MVEv and the incidence of AE will correspondingly increase. If these thresholds are exceeded only during major flooding in the Murray-Darling basin, outbreaks/epidemics of AE will remain a rare event. That is, as flooding in the Murray-Darling basin is correlated to major variations in the ENSO, the future incidence of AE will depend on the impact of the greenhouse effect on the ENSO.
The greenhouse effect, through changes to temperatures and the pattern of seasonal and geographical variation in rainfall, will alter the prevalence of mosquito vector populations in Australia. In turn, this may affect the potential for transmission of mosquito-borne diseases.
In the tropics, the potential for malaria reintroduction could be increased, and may extend to localities where the current climate is too erratic to support significant populations of vector Anopheles mosquitoes. The extended activity of Cx annulirostris and Ae vigilax might enhance the prospects for transmission of RRv and increase the incidence of EPA. Enzootic transmission of MVEv may also be enhanced and the geographical extent of activity enlarged. This may result in more frequent and intense activity near the southern limits of the enzootic zone in the Pilbara and Gascoyne Regions of Western Australia, and possibly in the Northern Territory and Queensland, and may correspondingly increase the incidence of AE.
In south-east Australia, the activity of vectors, particularly Cx annulirostris is likely to be increased, and the frequency of EPA may well be enhanced. The impacts on the infrequent outbreaks/epidemics of AE are more difficult to predict. If the altered conditions favour the formation of appropriate threshold associations of vectors and vertebrate hosts, the incidence of AE may increase. If these thresholds are exceeded only in those years of major flooding, the incidence of AE will depend largely on the impact of the greenhouse effect on the ENSO.
Anderson. S.G. (1952): Murray Valley encephalitis - epidemiological features. Med.J.Aust. 1, 97-100.
Anderson, S.G. and Eagle, M. (1953): Murray Valley encephalitis: the contrasting epidemiological picture in 1951 and 1952. Med.J.Aust., 1, 478-481.
Anon. (1987): Epidemiological features of Australian encephalitis. CDI 87/2, 12-14.
Bryan, J.H. and Russell. R.C. (1983): Australasian malaria vectors. Trans.R.Soc.Trop.Med.Hyg., 77, 278-279.
Bucens,M.(1982): Arbovirus infections diagnosed in Perth. 1981. pp. 171-178 In: Arbovirus Research in Australia, Proceedings 3rd Symposium, 15-17 February 1982. St George, T.D.and Kay, B.H.(eds. CSIRO, QIMR. Brisbane, 341pp.
Davies, F.G., Linthicum, K.J. and James, A.D. (1985): Rainfall and epizootic Rift Valley Fever. Bull.WHO, 63, 941-943.
Foo, L.C., Lim, T.W. and Fang, R. (1985): Rainfall, abundance of Aedes aegypti and dengue infection in Selangor, Malaysia. Southeast Asian J.Trop.Med.Pub.Hlth. 16, 560-568.
Forbes, J.A. (1978): Murray Valley encephalitis 1974, also the epidemic variance since 1914 and predisposing rainfall patterns. Aust.Med.Publ.Co., 20pp.
Grady, G.F., Maxfield. H.K., Hildreth, S.W., Timperi Jr, R.J., Gilfillan, R.F., Rosenau, B.J., Francy, D.B Calisher, C.H., Marcus, L.C. and Madoff, M.A. (1978): Eastern equine encephalitis in Massachusetts, 1957-1976. A prospective study centred upon analyses of mosquitoes. Am.J.Epidemiol. 107, 170-178.
Hawkes, R.A., Boughton. C.R., Naim, H.M. and Stallman, N.D. (1985): A major outbreak of epidemic polyarthritis in New South Wales during the summer of 1983/1984. Med.J.Aust., 7, 330-333.
Kay, B.H., Barker-Hudson, P., Stallman, N.D., Wiemers, M.A., Marks, E.N., Holt. P.J., Muscio, M. and Gomman, B.M. (1984a): Dengue fever, reappearance in northern Queensland after 26 years. Med.J.Aust., 140, 264-268.
Kay, B.H.. Barker-Hudson, P., Hapgood. G.D., McCurley, J.O., Lyons G.C. and Ives W. (1987): Aedes aegypti and dengue in the Townsville area. 1982-1985. Gen.Appl.Ent., 19, 2-10.
Kay, B.H., Fanning, I.D. and Carley, J.G. (1984b): The vector competence of Australian Culex annuluostris with Murray Valley encephalitis and Kunjin viruses. Aust.J.Exp.Biol.Med.Sci., 62, 641-650.
Liehne, P.F.S., Anderson, S., Stanley, N.F., Liehne, C.G. Wright, A.E., Chan. K.H.. Leivers, S., Britten D.K. and Hamilton, N.P. (1981): Isolation of Murray Valley encephalitis virus and other arboviruses in the Ord River Valley 1972-1976. Aust.J.Exp.Biol.Sci., 59, 347-356.
Liehne, C.G., Leivers, S., Stanley, N.F., Alpers, M.P., Paul, S., Liehne, P.F.S. and Chan, K.H.(1976): Ord River arboviruses - isolations from mosquitoes. Aust.J.Exp.Biol.Med.Sci., 54, 199-504.
Mahon, R.J. and Miethke, P.M. (1982): Anopheles farauti No.3, a hitherto unrecognized biolooical species of mosquito within the taxon A. farauti Laveran (Diptera: Culicidae). Trans.R.Soc.Trop.Med.Hyg., 76, 8-12.
Marshall, I.D. (1979): Epidemiolooy of Murray Valley encephalitis in eastern Australia - patterns of arbovirus activity and strategies of arbovirus survival. pp.47-53. In: Arbovirus Research in Australia. Proceedings 2nd Symposium, 17-19 July 1979. St George, T.D. and French, E.L. (eds.). CSIRO, QIMR, Brisbane, 169pp.
Marshall, I.D., Brown, B.K., Keith, K., Gard, G.P. and Thibos, E. (1982c): Variation in arbovirus infection rates in species of buds sampled in a serological survey during an encephalitis epidemic in the Murray Valley of South-eastern Australia, February 1974. Aust.J.Exp.Biol.Med.Sci., 60, 471-478.
Marshall, I.D., Thibos, E. and Clarke, K. (1982a): Species composition of mosquitoes collected in the Murray Valley of south-eastern Australia during an epidemic of arboviral encephalitis. Aust.J.Exp.Biol.Med.Sci., 60, 447-156.
Marshall, I.D., Woodroofe, G.M. and Hursch, S. (1982b): Viruses recovered from mosquitoes and wildlife serum collected in the Murray Valley of south-eastern Australia, February 1974, during an epidemic of encephalitis. Aust.J.Exp.Biol.Med.Sci., 60, 457-470.
McDonald, G. (1980): Population studies of Culex annulirostris Skuse and other mosquitoes (Diptera: Cullcidae) at Mildura in the Murray Valley of southern Australia. J.Aust.Entomol.Soc., 19, 37-40.
Miles, J.A R and Howes, D.W.(1953): Observations on virus encephalitis in South Australia. Med.J.Aust., 1, 7-12.
Mogi, M. (1983): Relationship between numbers of human Japanese encephalitis cases and summer meteorological conditions in Nagasaki, Japan. Am.J.Trop.Med Hyg., 32, 170-174.
Monath, T.P. (1980): Ch.6. Epidemiology. pp.239-312. In: St. Louis Encephalitis. Monath, T.P. (ed.). Am.Publ.Hlth.Assoc.
Murray. M.D. (1970): The spread of ephemeral fever of cattle during the 1967-68 Epizootic in Australia. Aust.Vet.J., 46, 77-82.
Newton, L.G. and Wheatley, C.H. (1970): The occurrence and spread of ephemeral fever of cattle in Queensland. Aust.Ver.J., 46, 561-568.
Nicholls, N. (1986): A method for predicting Murray Valley encephalitis in southeast Australia using the southern oscillation. Aust.J.Exp.Biol.Med.Sci, 64, 578-594.
Pittock,A.B.(1983): Recent climatic change in Australia: Implications for a CO2-warmed earth. Climatic Change, 5, 321-340.
Pittock, A.B. and Salinger, M.J. (1982): Towards regional scenarios for a CO2-warmed earth. Climatic Change, 4, 23-40.
Russell, R.C. (1986a): Seasonal activity and abundance of the arboviral vector Culex annulirostris Skuse near Echuca, Victoria. in the Murray Valley of southeastern Australia 1979-1985. Aust.J.Exp.Biol.Med.Sci., 64, 97-103.
Russell, R.C. (1986b): Seasonal abundance of mosquitoes in a native forest of the Murray Valley of Victoria, 1979-1985. J.Aust.Ent.Soc., 25, 235-240.
Russell, R.C. (1987): Age composition and overwintering of Culex annulirostris Skuse (Diptera: Culicidae) near Deniliquin, in the Mutray Valley of New South Wales. J.Aust.Ent.Soc., 26, 93-96.
Schlesinger, M.E. and Mitchell, J.F.B. (1987): Climate Model simulations of the equilibriurn climatic responses to increased carbon dioxide. Rev.Geophys., 25, 760-798.
Sellers, R.F. (1980): Weather, host and vector - their interplay in the spread of insect-borne animal virus diseases. J.Hyg.Camb., 85, 65-102.
Service, M.W.(1980): A guide to medical entomology. Macmillan & Co, London and Basingstoke, 226pp.
Stanley, N.F. (1982): Ch.32. Virus Diseases in Northern Australia. In: Viral Diseases in South-East Asia and the Western Pacific. Mackenzie, J.S. (ed.), pp.275-282. Academic Press, Sydney, New York, London, Paris, San Diego, San Francisco, Sao Paulo, Tokyo, Toronto, 751pp.
Wright, A.E., Anderson, S., Stanley, N.F., Liehne. P.F.S. and Britten, D.K. (1981): A preliminary investigation of the ecology of arboviruses in the Derby area of the Kimberley region, Western Australia. Aust.J.Exp.Biol.Med.Sci., 59, 357-367.
{kind=link}
{kind=link}