 Reproduced, with permission, from:
Longstreth, J. D., and J. Wiseman. 1989. The potential impact of climate change on patterns of infectious disease in the United States. In The potential effects of global climate change on the United States: Appendix G Health, ed. J. B. Smith and D. A. Tirpak for the Office of Policy, Planning, and Evaluation, U.S. Environmental Protection Agency. Washington, D.C.: U.S. Environmental Protection Agency.
Reproduced, with permission, from:
Longstreth, J. D., and J. Wiseman. 1989. The potential impact of climate change on patterns of infectious disease in the United States. In The potential effects of global climate change on the United States: Appendix G Health, ed. J. B. Smith and D. A. Tirpak for the Office of Policy, Planning, and Evaluation, U.S. Environmental Protection Agency. Washington, D.C.: U.S. Environmental Protection Agency.
Contract No. 68-01-7289
The United States Environmental Protection Agency (EPA) is concerned that these climatic changes will increase the morbidity and mortality of infectious disease in the United States. A two-day workshop was convened to address the questions relating to this issue, including the following: Which infectious diseases will become a greater health problem; what public health measures can be taken to lessen the potential impact of these diseases; and what research agenda should be established in regard to these issues.
The workshop decided that given the present levels of sanitation, immunization, and nutrition in the United States, malaria, dengue, yellow fever, and several skin diseases pose the greatest threat of increased morbidity and mortality.
The workshop also concluded that the decrease in the support of public health programs, particularly those in disease surveillance and vector abatement, create the greatest threat to effectively combating increased morbidity and mortality caused by climate change. It was also concluded that the development of multidisciplinary teams including experts from the fields of behavioral science, entomology, and epidemiology, need to be established in order to develop programs to effectively meet the changes in infectious disease patterns caused by climate change.
The climate[2] of the United States is expected to change due to an overall net global warming, which has been predicted to result from the increased global concentration of a number of "greenhouse gases" such as methane, carbon dioxide, and chlorofluorocarbons (CFCs) (EPA/UNEP, 1986; NAS, 1983). These gases are called greenhouse gases because they allow incoming solar radiation to pass through the earth's atmosphere but absorb outgoing infrared radiation thus resulting in warming within the confines of the atmosphere much as is seen within a greenhouse. Within the United States, it is expected that changes in temperature and rainfall will vary considerably by region, with some regions becoming colder and others warmer, as well as wetter or drier. Figure 1 presents one set of changes in temperature and rainfall, those predicted by the General Circulation Model (GCM) developed by NASA's Goddard Institute for Space Studies. Other GCM model predictions differ significantly from NASA's. While the quantitative and qualitative degree of regional climate change varies from model to model, it is relatively certain that temperature will increase by three to five degrees on average with a doubling in CO2. This paper attempts to discuss, based on available data, how infectious disease may vary with normal changes in climatic factors.
EPA has been requested to prepare a report for Congress on the potential adverse effects that might be seen in the United States due to the global warming. One of EPA's concerns is that, associated with changes in climate throughout various regions of the United States, there will be changes in the morbidity or mortality of infectious diseases[3], both those endemic in the United States and those which may be introduced from other locations. This report addresses EPA's concerns with regard to the infectious diseases that currently occur in the U.S. or that plausibly might be imported; the worldwide impact of climate change on infectious diseases is beyond the scope of this report. The document has two goals. One goal is to attempt to determine the likelihood (with attendant uncertainties) that increased morbidity and mortality from particular infectious diseases will occur in the U.S. owing to changes in climate, and the second goal is to identify the information necessary to reduce the uncertainty in these conclusions.
It has been many decades since infectious diseases were the major causes of mortality in the United States, as they are now in large parts of the world. Figure 2 compares the 10 leading causes of death in the United States for 1900 and 1981. The leading causes of mortality at the turn of the century were (Mauser and Kramer, 1985) pneumonia and influenza, with tuberculosis second. By 1980, pneumonia and influenza were significantly reduced in importance and tuberculosis was not one of the ten leading causes of death. The differences in the leading causes of death in the United States between 1900 and 1980 are principally due to improved sanitation, housing, nutrition, immunization programs, and treatment. It seems unlikely that climate change will cause a return of these diseases as important causes of mortality.
Nevertheless, certain infectious diseases are not necessarily mitigated by improvements in sanitation, nutrition, immunization and treatment. These diseases present the greatest public health threat either because there are no vaccines or treatments currently available, because the infectious agents are not significantly impacted by sanitation or nutrition and/or because there may be significant portions of the United States population who have substandard access to such controlling factors. For the most part, diseases with these characteristics appear to be those that are vector-borne[4]. Thus this report focuses on this subset of infectious diseases in order to evaluate whether these diseases are likely to show changes of incidence within the United States following changes in climate. This evaluation will be based principally on information derived from five examples of vector-borne diseases, some of which currently exist in the United States and some of which either once existed in the U.S. and/or exist in locations near its borders. This subset of infectious diseases was chosen because a significant amount of information suggests that the prevalence and distribution of the vectors of these diseases can be affected by climatic variables, which in turn suggests that changes in climatic variables could change the prevalence and distribution not only of vectors but, most importantly, of the agents they carry.
There are at least two ways in which climatic change may impact vector-borne diseases. One mechanism of change is brought about by the direct impact of climatic change on the agent, vector, or host. For instance, changes in temperature, rainfall, humidity, or storm patterns that directly impact the multiplication or differentiation rate of the vector or the agent, increase the biting rate of the vector, or increase the amount of time that the host is exposed to the vector, would be considered direct impacts of climate change. A second mechanism of change is brought about by the indirect impacts of climate. In this category, climate influences some parameter that is important to vector spread or survival, such as the type of agriculture or the species of trees in a forest; this, in turn, changes the relationship between the parasite, vector, and host. Both types of impacts are considered in this report; clearly, however, the direct impacts are easier to evaluate than the indirect ones.
The report[5] first evaluates what is known about the impact of climatic factors on five vector-borne diseases: Lyme disease, Rocky Mountain spotted fever, malaria, dengue fever, and viral encephalitis. A discussion of several additional diseases determined by the workshop participants to be important is then presented. The information from these sections is analyzed for what can be generically stated about the effects of climate on agents, vectors and hosts. The final section presents the summary and policy implications.
LYME DISEASE
Lyme disease, initially recognized in Lyme, Connecticut, in 1975, is caused by a spirochete, Borrelia burgdorferi, and is transmitted by ticks of the Ixodes ricinus complex[6]. It is an inflammatory disease, characterized by a distinctive skin lesion, erythema chronicum migran (ECM), systemic symptoms (profound fatigue, fever, chills, headache, and backache), polyarthritis, and cardiac and neurologic abnormalities which occur in varying combinations (Habicht et al., 1987). Symptoms are acute, lasting several weeks for most patients. Treatment with antibiotics is available and eventually complete recovery ensues (Braunwald et al., 1988).
The life cycle of the tick vector normally spans two years. Eggs are laid in the spring to hatch a month later into the larval form. During the first summer, the larva feed once on the blood of the host, then enter a resting stage with the onset of cold weather. The next spring, the larvae molts to become a nymph, which again attaches itself to an animal host. The nymph stage is primarily responsible for disease transmission (Steere, 1979). At the end of the summer, the nymphs molt into adults. They can be found in brush about one meter above the ground, where they easily attach to larger mammals (Habicht et al., 1987). Lyme disease is usually contracted between May 1 and November 30. The majority of cases are acquired in June and July (Steere, 1983b). This time frame corresponds with the peak questing period of Ixodes dammini,which for nymphs is May through July.
The principal hosts for juvenile and mature forms of I. dammini are the white-footed mouse (Peromyscus leucopus) and the white-tailed deer (Odocoileus virginianus), respectively. In addition, approximately 80 species of birds, mammals, and lizards have been identified as hosts of I. pacificus. It appears that lizards and Columbian black-tailed deer are the most important host of immature and adult I. pacificus, respectively (Westrom et al., 1985; Lane and Burgdorfer, 1986). The major importance of migrating birds as hosts is that they facilitate the movement of infected vectors over a large geographic radius, thereby substantially enlarging the geographic range of the vector and the agent (Hoogstraal et al., 1963; Spielman et al., 1985). In areas on the North Atlantic coastline, 80-90% of Ixodes ticks have B. burgdorferi in their gastrointestinal tracts. This rate contrasts with only a 3% rate of infection of I. pacificus on the West Coast. These rates correlate with the relative prevalence of the disease in the two areas.
Although Lyme disease has been reported from over 25 states, it has four major foci in the United States (Figure 3). It has also been reported in Germany, Switzerland, France, and Austria. In 1975, 59 cases were recorded in Connecticut; in 1985, the number had climbed to 863 cases (Habicht et al., 1987). Similar statistics are reported from New Jersey, with incidence increasing from 14 cases in 1980 to 39 in 1981 to 56 in 1982 (Bowen et al., 1984). Age-specific attack rates show that the risk of contracting Lyme disease is about equal in all age groups through age 50. After age 50, the risk lessens, probably because older persons are less likely to be present in tick-infested areas (Steere, 1983a). Better reporting may contribute to the increase in incidence statistics.
Lyme disease appears to occur wherever the vector is abundant. The presence of the vector in the northeast seems to depend on the presence of the white-tailed deer (Main et al., 1982). The range of the northeastern subspecies (O.v. borealis) is the known range of the vector I. dammini (Spielman et al., 1985), although formal proof of a cause and effect relationship is lacking (Spielman et al., 1984). Tick abundance may also depend upon environmental factors such as temperature, humidity, vegetation, and physiographic features of the environment (Spielman et al., 1985). Previous attempts to determine the abundance of I. dammini solely by climate (McEnroe, 1977) failed in that it ignored the distribution of the deer host and the presence of the tick in Wisconsin, which is a different climatic region (Spielman et al., 1985).
ROCKY MOUNTAIN SPOTTED FEVER
Rocky Mountain spotted fever[7] (RMSF) (also known as tick-typhus) is caused by the small, gram-negative, coccobacillus Rickettsiae rickettsii. The disease is characterized by fever, chills, headache and rash. Antibiotics are available for treatment, and vaccinations are available for those at greatest risk for developing the disease (Braunwald et al., 1988).
The two principal vectors of RMSF are the dog tick, Dermacentor variabilis, the distribution of which is shown in Figure 4; the wood tick Dermacentor andersoni, which is found mainly in the Rocky Mountain states; and Dermacentor occidentales which is found in parts of the western United States. Infection is maintained in ticks by transovarial transmission and by infectious feeding. The opportunity for a tick to acquire infectious rickettsiae is limited to the short period (3-4 days) in the life of a susceptible animal when the level of circulating R. rickettsii in the blood of the host is high enough for the tick to receive an infective dose[8] (McDade and Newhouse, 1986).
No data were found concerning the impact of environmental temperatures on the infectivity of R. rickettsii; however, data on R. mooseri, the agent for murine typhus, indicates that ambient temperature has a profound effect upon rickettsial growth in fleas, as well as the survival of fleas themselves. At 18deg.C, the rickettsial content of the fleas was below detectable levels for at least ten days and remained low throughout, whereas at 24deg. or 30deg.C, the rickettsial titer was consistently two or three times greater. In addition, if, after six days, the fleas were transferred from an environment of 18deg.C to one of 24deg. or 30deg.C, the rickettsial growth increased by two or three logs within one week (Farhang and Traub, 1985). The coccobacillus is heat tolerant.
Infection occurs in a large number of animal hosts. For example, in Maryland and Virginia, antibodies were found in 15 different mammals and 18 different types of birds (Bozeman et al., 1967; Sonenshine and Clifford, 1973). This large spectrum of animal reservoirs, the fact that several tick species are naturally infected with R. rickettsii (McDade and Newhouse, 1986), and man's role as an incidental host in the natural cycle makes eradication of RMSF unlikely. This is especially true since the United States has no tick control programs (Dr. Dan Haile, USDA; personal communication[9].
Climatic, ecologic, and geophysical variables influence the timing of outbreaks. RMSF is a seasonal disease occurring in the warm periods that coincide with increased tick activity. In the west, the peak time is usually the spring and in the east the peak is from May through September.
Data for the United States population as a whole for 1970-1980 showed more than a doubling in incidence for the first half of this decade, going from less than 0.2 per 100,000 in 1970 to greater than 0.5 per 100,000 in 1977 (CDC, 1986f). The higher rate was maintained until about 1983, falling to about 0.4 in 1984. Whether these changes were due to patterns of land use, microenvironmental changes, or other factors could not be determined from the available data
Although the highest incidence of the disease is in children aged 5-9,[10] the highest mortality rates occur in the 40-59 age group[11] (D'Angelo et al., 1982). The fatality rate is about 15 to 20% in the absence of specific therapy; with prompt recognition and treatment, death is uncommon, yet between 4 and 6% of cases reported in the United States during recent years have died (APHA, 1980). Figure 5 shows the geographic distribution of the disease for 1982.
Prevalence of RMSF has been linked to natural vegetation, which reflects regional climatic conditions. The density of the principal tick vector also varies with the natural vegetation of the area The highest incidence of RMSF has been associated with oak-hickory-pine forests. Within this general range, highest incidence is associated with drier, mesic forest types (Sonenshine et al., 1972)[12].
MALARIA
Malaria affects a large geographic area. Over the past 10-15 years, the prevalence and geographic distribution of malaria worldwide has increased slowly but steadily, sometimes in small foci, and sometimes over whole sub-continents. Its recent worldwide increase is due mainly to mosquito resistance to pesticides, breakdown of control efforts, migration of the vectors, and irrigation. However, its spread is also environmentally related. Man's activities, including agriculture and road building, have created better habitats (e.g., more still water) and thus contribute to the spread of malaria into several areas where it was not previously present (De Zulueta 1980).
Four protozoan agents potentially cause malaria in humans: Plasmodium vivax, P. malariae, P. ovale, and P. falciparum. The most serious form of malaria is induced by P. falciparum and presents a very varied clinical picture including fever, chills, sweats, and headache. It may progress to liver damage, coagulation defects, shock, renal and liver failure, acute encephalitis and coma. Case fatality among untreated children and nonimmune adults exceeds 10%. Malaria induced by the other Plasmodium sp. is generally not life threatening except in the very young, the very old, or those with concurrent disease. The less severe forms of malaria are characterized by an initial feeling of malaise, usually accompanied by headache and nausea and ending with profuse sweating. This pattern of symptoms occurs in cycles which appear daily, every other day, or every third day. Treatment of acute attack can be accomplished with quinine or chloroquinine (Braunwald et al., 1988).
Plasmodia are transmitted to humans via the bite of anopheline mosquitoes. In the mosquito, plasmodia go through a complicated life cycle involving a number of differentiation stages. P. vivax and P. malariae require environmental temperatures of at least 15deg.C for their development within the anopheline mosquito. P. falciparum requires temperatures of at least 17deg. or 18deg. C for its development to take place (Macdonald, 1957). With gradual increases in global temperatures since the last ice age (+8.0-9.5deg.) transmission has migrated from Africa to southern Europe.
A number of different anopheline mosquitoes can serve as vectors for the malaria agents. In the United States, these include Anopheles quadrimaculatus and A. freebornei. In addition, Plasmodium species and isolates are known to adapt in order to be able to develop in particular Anopheles species or subspecies (WHO, 1987).
Malaria was once a significant health problem in the southern and western United States (Faust, 1938). Figure 6 illustrates mortality due to malaria in the south for the period 1925-35. With the realization of the role played by the mosquito in malaria transmission, breeding of the mosquito was controlled by better drainage and ambient spraying (Williams, 1938; Williams, 1937). Resistance of the vector to pesticides can make present-day control programs more difficult. Both P. falciparum and P. vivax were present in the United States.
Indigenous transmission of malaria within the United States, although rare, is still possible. Some or all of the 27 reported cases of P. vivax that occurred in San Diego County in June-September 1986 occurred through indigenous transmission. A competent vector, Anopheles freeborni, is found in the southern California. In addition, there have been reports of local transmission among Punjabi immigrants in the Sacramento Valley. This represent the largest outbreak of malaria in the United States since 1952 (Brunetti et al., 1954; CDC, 1986a,f).
With a large influx of immigrants, sufficiently large amounts of the malaria parasite could be introduced into the United States in the presence of competent vectors. These vectors include An. freeborni in California and the states that border Mexico, as well as A. quadrimaculatus[13] in the entire Southeast during the warmer months (CDC, 1986a). As the climate becomes warmer, it may be expected that these vectors will increase in their geographic range throughout the entire Southeast during the warmer months (CDC, 1986a), and that An. quadrimaculatus will be present for a larger part of each year. In addition, raising the ambient temperature could provide conditions more favorable to the replication of the various Plasmodium sp. within a competent vector. If more irrigation is required in areas with increasing temperature and adequate drainage is not concurrently supplied, more mosquitoes could breed (Gratz, 1973). Longer seasons for exposure of the host human to infected vectors could also result.
DENGUE FEVER
Dengue is a mosquito-borne virus with four serotypes (Dengue 1-4). It is classified as a Group B arbovirus and is antigenically related to St. Louis encephalitis, yellow fever, Japanese B encephalitis, and other viruses. Classic dengue fever, which usually is not fatal, is characterized by the abrupt onset of fever and generalized body aching as well as severe headache and retro-orbital pain. Dengue fever can be complicated by dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS), both of which can be fatal, particularly in young children.
The principal vector for dengue is the Aedes aegypti mosquito, whose distribution in the United States is shown in Figure 7. A potential vector, Aedes triseriatus, is found throughout the eastern half of the United States. In addition, Aedes albopictus, a vector for dengue, originally of northern Asiatic origin, and resistant to winter freezing temperatures, has now been found in the United States. Aedes albopictus[14] has both rural and urban habitats and is a competent vector for dengue (Anon., 1987a). The distribution of A. albopictus in the U.S. is shown in Figure 8.
Dengue is not presently endemic in the continental United States. However, epidemics have occurred in the past[15]. The last such large epidemic occurred in Texas in 1945 (Anon., 1987a). Nine cases of indigenous transmission of dengue within the United States were reported from Texas in 1986 (Reiter, 1988).
Although dengue is not currently indigenously transmitted in the continental United States, it is a problem in Puerto Rico and also several localities that provide large numbers of immigrants to the United States. For example, 2,371 cases of dengue (DEN-1, 2 and 4) were reported from Puerto Rico in 1985 (Anon., 1987b). In addition, dengue had not been reported in Cuba for 30 years, when a DEN-1 epidemic occurred in 1977. This was followed in 1981 by an epidemic of DHF caused by DEN-2, which resulted in at least 158 deaths (CDC, 1981). Similarly, in 1978, Mexico reported its first cases of dengue in many years. The disease became endemic in most coastal areas of Mexico, and in 1984 cases of DHF were reported (CDC, 1986e). The spread of dengue and DHF in these countries is following a pattern similar to that found in Southeast Asia, where DHF is presently a leading cause of hospitalization and death of children.
Dengue is frequently introduced into the United States by people who have traveled abroad, who return to areas containing a competent vector for dengue (illustrated in Table 1 for 1982). Although travelers importing dengue virus into the United States have not introduced a large amount of the virus into a small geographic area, large numbers of immigrants from dengue endemic areas tend to settle in small geographic areas. A competent vector for dengue may be present in these areas.
Specific experiments have been conducted on the effect of temperature[16] on the ability of Aedes aegypti to transmit DEN-2 virus. These experiments showed that the DEN-2 virus was transmitted by A. aegypti only if the mosquitoes were kept at 30deg.C. The required extrinsic incubation period was shortened if the temperature was increased to 32deg.C and 35deg.C (Watts et al., 1987). This pattern of temperature and vector efficiency parallels the climatic pattern of DHF outbreaks in Bangkok, Thailand, where case rates rise during the hot season (with daily mean temperatures of 28deg.-30deg.C ) and decrease during the cool season (with daily mean temperatures of 25deg.-28deg.C) (Watts et al., 1987).
ARBOVIRUS-RELATED ENCEPHALITIS
Arthropod-borne viruses (arboviruses) are associated with several major clinical syndromes[17], including encephalitis. Arbovirus infections were responsible for 65% of diagnosed encephalitis cases reported to CDC between 1969 and 1979 (Shope, 1980). At least 18 arboviruses cause encephalitis; of these, at least 7 are present in the United States, including western equine encephalitis (WEE), St. Louis encephalitis (SLE), eastern equine encephalitis (EEE), California encephalitis (CE), La Crosse encephalitis, Powassan encephalitis, and Venezuelan equine encephalitis (VEE).
The effects of infections with these viruses range from a mild influenza-like syndrome to central nervous system (CNS) disease, which can be fatal. Encephalitis is characterized by acute febrile illness (oral temperature >= 100deg.F), with signs of brain parenchymal inflammation that include one or more signs of decreased level of consciousness (confusion, disorientation, delirium, lethargy, stupor, coma) and/or objective signs of neurologic dysfunction (convulsion, cranial nerve palsy, dysarthria, rigidity, paresis, paralysis, abnormal reflexes, tremor, etc.). Meningeal irritation is often seen in encephalitis patients; symptoms may include aseptic meningitis (acute febrile illness and signs of meningitis, such as stiff neck) and febrile headache (Monath, 1980). Delayed or latent CNS sequelae in children who have had encephalitis is also a severe complication (Finley and Longshore, 1958).
Outbreaks of encephalitis caused by the different viruses are normally limited to specific geographic locations and seasons, including seasons in which arthropod breeding and feeding occur. Most cases of encephalitis occur in late summer/early fall when mosquitoes, the primary vectors, are prevalent. In addition, infection depends on certain variables, including the species of mosquitoes that are susceptible to a specific virus, and the viral concentration in the susceptible vertebrate host's blood. More than 100,000 infectious units per milliliter are usually required in order for virulent strains to infect mosquitoes. An extrinsic incubation (EI) period, i.e., the interval between ingestion of the virus and subsequent transmission through biting, of 4 days to 2 weeks at summer temperatures is normally required, before the virus is fully multiplied and can be transmitted to a new host.
Many of these variables are affected by climatic change. For example, environmental temperature affects the EI (the species discussed here are active at temperatures of 13deg.-35deg.C). Increased temperature can decrease the EI, thereby quickening the transmission process and promoting epidemic disease. Moisture, present as rainfall or irrigation, affects the growth of plant life for feeding of host animals, and the presence of insect breeding sites. Changes in these environmental conditions affect the ability of the vectors to transmit the virus effectively. Finally, viruses carried by the same vectors appear to occur in the same climatic conditions and geographical distributions. It would follow, then, that the effective spread of the viruses, and usual subsequent epidemic disease, is dependent upon optimal environmental conditions in which the vectors may breed, feed, and transmit the viruses. Those conditions, wet or dry, mild or warm climates, vary with the particular species of mosquito vector.
WEE is transmitted in a mosquito-bird cycle[18] by the vector Culex tarsalis. C. tarsalis mosquitoes breed in groundwater ponds and abound in irrigated areas of California, Texas, and other parts of south-central and southwestern United States, where SLE occurs endemically (Shope, 1980). SLE accounts for occasional small outbreaks each year in humans and for periodic large epidemics. Warm weather is required to complete the extrinsic incubation of the virus in the mosquito. As the C. tarsalis naturally feeds on large vertebrates, equine and human cases occur annually. WEE, which can be severe but is not normally fatal, is seen primarily in children under 2 years of age, causing retardation, seizures and spasticity (Johnson, 1977).
Rainfall[19] strongly affects the numbers of infected individuals, since C. tarsalis breed mainly in ground pools and irrigated ponds. Environmental temperature also affects the activity of the arbovirus. The maximum temperature permissible for the WEE vector to transmit the virus effectively was <= 25deg.C. Above 32deg.C, virus transmission rates rapidly decreased (Hardy et at., 1980; Kramer et al., 1983). A study of EI temperatures of C. tarsalis showed decreased vector competence after two to three weeks of EI at 32deg.C as compared with vector competence after EI at 18deg. or 25deg.C. The high temperature did not, however, affect preexisting infection (Kramer et al., 1983). Studies also demonstrate lower effectiveness in WEE virus transmission to humans at higher ambient temperatures (Hardy et al., 1980; Kramer et al., 1983). The cooler temperatures at which the WEE virus is better able to replicate allow for epidemic disease much earlier in the summer, and eventually much farther north in cooler climates later in the season (Shope, 1980).
SLE is the most geographically widespread arbovirus in the United States and the most common cause of epidemic arbovirus encephalitis in humans (Johnson, 1977). The C. tarsalis mosquito transmits the virus in the western United States and C. nigripalpus in Florida in the same manner in which WEE is transmitted. The two viruses carried by these vectors are therefore affected in the same way by climatic change. Thus, the vector and the infection are prevalent in areas and seasons of cooler climate and high precipitation.
SLE[20] in humans has been reported throughout most of the United States; however, the first reported Canadian case in humans occurred in 1975 (Spence et al., 1977). During that same year, a widespread epidemic occurred, with over 2,000 reported cases and 171 deaths, predominantly throughout the eastern two-thirds of the United States. The epidemic was primarily between late June and October. During that year, precipitation was below the long-term (1941-1970) average in June and July, and temperatures exceeded the normal in May. Figure 9 shows the percentage of cases of arboviral encephalitis by month of onset and the etiology reported to the CDC between 1955 and 1971. SLE encephalitis occurred mainly in the late summer and tended to follow those due to CE and WEE, but tended to precede cases of EEE (Monath, 1980). Nearly all epidemics have shown that the incidence of SLE is 5-40 times higher in persons over 60 years than those in the 0-9 age group (Monath, 1980).
Mosquitoes belonging to the Culex pipiens complex (C.p. pipiens, C.p. quinquefasciatus, and C. nigripalpus) are the principal vectors for SLE in urban areas. These species breed in polluted water of high organic content; therefore, these species are suited to urban and suburban areas (Monath, 1980; Shope, 1980), unlike the primarily rural C. tarsalis. In contrast to the effect of rainfall on C. tarsalis, C. pipiens epidemics occur more often in times of drought and high temperatures. The lack of rainfall results in poorly draining and stagnant water, providing increased breeding grounds for the vector (Monath, 1980; Shope, 1980; Johnson, 1977). The year of the first major epidemic in the United States, 1933, was the driest year recorded since 1837 (Shope, 1980).
High temperatures favor virus transmission for C. pipiens and C. tarsalis by decreasing the EI time, as well as the time required for larval maturation and development of viral infectivity (Monath, 1980). Studies of the relationships between incubation time of SLE virus in C. pipiens and mean temperature found that daily exposures to increased temperatures from a constant of 25deg.C decreased incubation time, thereby increasing effectiveness of viral transmission (Hardy et al., 1980). In addition, a review of all the arbovirus encephalitis cases found in the United States showed that most WEE outbreaks have occurred at or above the 70 F June isotherm, whereas most SLE cases have occurred in warmer latitudes at or below the June isotherm (Hess, 1963).
Eastern equine encephalitis (EEE) is transmitted in a mosquito-bird cycle by Cuiseta melanura and is present in the freshwater marshes along the Gulf and Atlantic coasts, including the Great Lakes region (Johnson, 1977; Shope, 1980). The vector mosquitoes do not feed on large vertebrates; therefore, infection in humans is very rare. However, climatic changes altering the conditions of the wetlands, such as rainfall, could introduce changes in mosquito breeding or types of susceptible birds. The virus might then spill over to a species of mammal-feeding mosquitoes, most likely Aedes or other Coque tidia, via infected birds (Shope, 1980). This is the only route of epidemic transmission of EEE to horses and to humans, in whom infection is rare, but fatal in 60% of the cases (Johnson, 1977). EEE affects primarily children, and death usually occurs in the first 2-5 days after onset, characterized by periorbital edema during the acute disease (Johnson, 1977).
The La Crosse strain of California encephalitis virus causes mild to severe cases of encephalitis in children ranging from a benign aseptic meningitis to a severe, but rarely fatal, form of encephalitis (Johnson, 1977). There is little information on the effect of climate on the occurrence of the disease. The virus is widely distributed in the Midwest and the middle Atlantic and Appalachian states (Shope, 1980), and is transmitted by Aedes triseriatus mosquitoes, which reside in woodland treeholes and feed on small woodland mammals. A. triseriatus transmits the virus transovarially,[21] (Shope, 1980) as well as venereally through infected seminal fluid (Thompson, 1978), and thus the adult progeny can then transmit the infection by bite upon adult emergence (Shope, 1980). This vertical transmission cycle allows for human La Crosse infection during early summer, and is amplified by infection and viremia in small mammals. The mosquito has a very limited flight range, so humans are infected only when exposed to the woods or to old discarded tires, which serve as "synthetic treeholes" (Johnson, 1977).
In the attempt to predict what the potential impacts of climate change might be on infectious diseases, we decided that the analysis would be facilitated through the development of a conceptual model that looked at the interrelationship between climate and the elements necessary for a vector-borne disease to occur. Accordingly, given below are brief summaries of information drawn from the five diseases discussed above, which apply to the three elements of a vector-borne disease: the agent, the vector, and the host (or hosts).
THE AGENT
The agent in a vector-borne disease may be a virus, protozoa, bacteria, or helminth. Infectious agents are transmitted to their hosts by vectors. The role of the vector may not only be one of transport, but may also be required for completion of the lifecycle of the agent. As indicated above, there are two potential concerns regarding the impact of climate change on disease agents: one is that climate change will allow the establishment of new agents in the United States; the other is that agents that are presently endemic will flourish, thereby favoring either enlargement of the geographic area where they are found or greater infectivity in the current geographic area.
Agents Not Currently Present in the U.S.
Malaria and dengue are two vector-borne diseases not currently established in the United States, although they are introduced regularly and have been established in the United States in the past. Two questions must be considered in evaluating whether the establishment[22] of new infectious diseases in the United States is possible. The first is whether the agent is currently present or is likely to become present (by virtue of immigration, etc.) within the geographic range of a competent vector; the second is whether conditions are or will become favorable to the development and spread of the agent. The minimum amount of agent in a geographic area needed to assure transmission of the disease varies with each specific disease.
Although from time to time infectious diseases may be introduced into the United States by the return of travelers, generally the geographic distribution of these individuals is sufficiently broad that the critical mass of the agent necessary for establishment is unlikely to occur. For malaria, climatic conditions do not favor the differentiation of the agent within the vectors which are present.
Currently, the two most likely sources of introduction of a disease[23] into the United States are via legal immigrants and refugees (principally from Asia) and via migrant workers and illegal immigrants. Between 1975 and 1981, 0.5 million Asian refugees settled in the United States. Roughly one third of that population lives in California, one third lives in the states of Texas, Washington, Pennsylvania, Illinois, Minnesota, Virginia and Oregon; and one third have settled throughout the remaining states (Cantanzaro, 1982). If immigrants continue to congregate in nearby areas, a critical mass could be approached. In addition to these refugees, between 1977 and 1985, the largest number of immigrants admitted to the United States have been from Asia (INS, 1985). Of the 50 metropolitan statistical areas specifically listed by the Immigration and Naturalization Service as areas where immigrants intended to reside, 14 are areas presently known to be infested with Aedes aegypti (INS, 1985; CDC, 1986e,f). The fact that a second vector, Aedes albopictus, has become established in northern, as well as southern states (CDC, 1986b,d), broadens the area in which a dengue virus could become established given appropriate conditions.
Another source for the introduction of new infectious agents into the U.S. is via migrant workers (border commuters) and illegal immigration. It is estimated that between 4.5 and 12 million illegal aliens reside in the United States (Bos, 1984), as well as another 1.3 million seasonal workers and border commuters (Corwin, 1982). It is assumed that these individuals carry the parasites commonly found in their home countries. In addition to dengue and malaria, large percentages of immigrants and refugees are infected with intestinal parasites (15% to 78%), positive tuberculin test (49% to 55%) (Arfaa, 1981), and hepatitis B (12% to 36%) (CDC, 1979a,b; 1980c,e; Barry, 1983; McGlynn, 1985; Cantanzaro, 1982; Jones et al., 1980).
Medical screening of immigrants and refugees does not include testing for arbovirus, malaria, or most parasitic diseases. The CDC guidelines for medical examination of non-United States citizens seeking entry into the United States do not include such diseases as one of the "dangerous contagious diseases."[24] Nevertheless, it is estimated that within a single refugee detention center, several cases of dengue are identified each year, with an occasional large outbreak. There are also confirmed cases of malaria each year.
This section has illustrated that sufficiently large amounts of parasites are introduced into the country, making outbreaks of diseases not previously endemic to the United States possible. Although other factors also need to be present, the first element of the infectious disease relationship is presence of the parasite.
Agents Currently Found in the U.S.
For agents of diseases such as RMSF, Lyme, arbovirus-transmitted encephalitis, plague, and other endemic agents, the critical question is whether changes in climate will either enlarge the area in which they are present or increase the number of cases seen in the endemic areas. With regard to the role of the agent, is it likely that replication of the agent in vectors (or hosts) will be more favored under new climatic conditions? If the answer to this question is yes, then this could result either in larger amounts of agent per vector, a shorter time for the agents to reach infectivity within the vector, or a wider geographic area that supports development of infectious agents in the vector. Given below are brief summaries of what was found in the literature relevant to assessing these questions. Clearly, much is not known. (A discussion of additional data needs is presented in the Conclusions and Recommendations section.)
Lyme disease currently has four major foci in the United States. The climatic conditions within the foci are dissimilar and the transmission of the disease seems to be more dependent on the presence of infected ticks and their principal hosts, the white-footed mouse and the white-tailed deer, than on the competence of the agent within the vector. No data were found to address whether climatic factors can affect the reproductive and/or developmental processes of this spirochete within its tick vector. However, the incidence of the disease is increasing.
The incidence of Rocky Mountain Spotted Fever in the United States doubled in the decade of 1970 to 1980, from less than 0.2 per 100,000 to more than 0.5 per 100,000; however, by 1984 the rate was 0.4 per 100,000. These are nationwide statistics and it is unclear what they reflect in terms of the agent. Clearly the agent is present; however, very little information was found upon which to base an assessment of how climate change could affect its distribution or infectivity. If R. rickettsia, the agent for RMSF, behaves like R. mooseri, the agent for tick typhus, then one would expect to see increased rickettsial titers in ticks under warmer conditions; however, as discussed below, warmer temperatures could disrupt diapause in the vector life cycle, which could adversely affect tick survival or rickettsial cycles.
THE VECTOR/ANIMAL HOST
Vector-borne diseases were selected for study in this review principally because the greatest amount of information found about the relationship of climatic variables to infectious diseases was found for this subset of infectious diseases, and the judgment was made that the high levels of nutrition, health care, and hygiene found in the U.S. were likely to prevent a significant impact of climate changes on other more common bacterial and viral diseases. The discussion that follows focuses principally on the impacts that climatic change could potentially have on the tick and the mosquito. In addition, this section includes information on the impact of climatic variables on intermediate vertebrate hosts.
The introduction of a vector into a disease process means that there are now three places (agent, vector/intermediate host, and human host) that require attention to control a disease, instead of the standard two (agent and host). Because of this intermediate step between the agent and the host, control of these diseases becomes more difficult than control of diseases such as pneumonia or tuberculosis, which are induced directly by the agent in the host.
The two most important factors related to a vector's transmission of disease are the geographic range (both in distance and in amount of time during the year that the vector is present) and the vector's rate of infectivity by any parasite. The geographic distribution of tick vectors in the United States is determined by a complex set of factors which are both geographic and climatic in nature. For example, D. andersoni, the vector of Rocky Mountain spotted fever (RMSF) in the western part of the United States, is associated with "shallow soils, moderate shrub cover, exposed rock, steep slopes, numerous pines, log litter" and decreasing grass cover (McDade and Newhouse, 1986). Those same factors influence the abundance of the vertebrate host for the larval tick.
Temperature and relative humidity are also important in the vector's spatial distribution. The relative humidity must be high enough to prevent desiccation of ticks and their eggs, and the ambient temperature must be high enough (app. 20deg.C) to allow the life cycle of the vector to be completed. However, winter soil temperatures must be low enough to release the adult ticks from their diapause, a period of lowered metabolism, that prevents their feeding in the fall (Wilkinson, 1967; McDade and Newhouse, 1986). This appears to be less than 5deg.C. Shorter survival rates have been observed at 75% relative humidity than at 85-95%, although the optimum relative humidity for vector reproduction and survival has not yet been determined (Wilkinson, 1967).
The difference in the timing of onset of disease, as contrasted by the pattern of RMSF in Virginia, Massachusetts, and Nova Scotia, may be explained by the direct or indirect effect of environmental changes. For example, a change in the temperature isotherms in the various areas could have modified a vector's life cycle, or a change in the composition of the flora and/or fauna could have resulted in the introduction of an additional vector whose life cycle differs temporally from the original vector so that maturing and overwintering ticks overlap considerably in terms of the time of activity. The definitive combination of factors that are conducive to the continued geographic spread of the vector has not yet been determined (Garvie et al., 1978).
The impact of climatic factors on mosquitoes serving as vectors has been studied for encephalitis virus infections. Transmission of arboviruses by mosquitoes is influenced by a number of factors. Among these are population levels, biting habits, and the intrinsic factors affecting the actual biological transmission of the specific agents. These variables are usually very closely related to climatic change, especially temperature and rainfall, which in turn determine the geographical distribution of the individual species and the viruses for which they are specific (specific viruses are carried only by specific susceptible vectors).
The population levels and biting habits of virus-specific mosquitoes vary according to climate, genetic determination, and abundance of hosts. In species which undergo diapause[25] and are thus capable of overwintering, the population levels[26] vary throughout the year. Viral activity is usually increased during the active summer months and decreased during winter months because of reduced feeding levels related to the onset of diapause. Diapause is induced in late-stage larvae and pupae by a combination of shortened photoperiod and cooler temperatures, and thus varies temporally with latitude (Bailey, 1982). In a study of Culex pipiens, indigenous to temperate climates and capable of overwintering, and Culex quinquefasciatus, found in pantropical regions (Florida) and incapable of hibernation, it was found that although both species develop fat reserves on a sugar diet, only C. pipiens within this complex of species underwent the additional physiologic changes associated with diapause in response to exposure to shortened photoperiod and decreased temperatures. The C.p. quinquefasciatus is apparently unable to utilize its fat reserves or decrease its activity in order to survive temperate-zone winters. The C.p. pipiens, on the other hand, showed signs of overall decreased activity, including reduced blood-feeding[27] (Eldridge, 1968).
In addition to influencing population levels and biting habits, climate variables strongly influence the capability and rate of transmission in vectors that undergo biologic transmission. Effective biologic transmission (or extrinsic incubation, EI) depends on several variables such as the infectivity threshold, virus multiplication rates, and susceptibility and dissemination barriers, many of which are temperature-dependent.
The EI period is the time required after ingestion of an infective blood-meal, during which the viral multiplication takes place and before the virus can be transmitted orally by the vector (Hardy et al., 1983). The specific time varies for each virus and mosquito, and the length of time the mosquito must be able to survive after ingestion in order to transmit the virus can be determined from it. Longer survival rates of mosquito populations, then, are required to maintain cycles of viruses with long EI periods (Hardy et al., 1983). Viral multiplication rates vary directly with temperature, as shown in a study of vector capability of Aedes aegypti for California encephalitis virus and dengue viruses at various temperatures. A. aegypti is indigenous to warmer climates and is a highly effective vector of dengue, a tropical arbovirus, but is an ineffective vector of CE[28], which is endemic at latitudes where frigid conditions prevail. At a range of temperatures, the EI period of DEN2 virus increased from 6-13 days with a decrease in temperature from 90deg.-75deg.F. Transmission of the Yukon strain of CE was very ineffective, with EI periods of 3 weeks at 80deg.F, and 4 weeks at 55deg.F (McLean, 1977). In a similar study of A. aegypti, viral replication of Yukon and Norway strains of CE and MVE (found in moderate climates) was seen in its full viable range (13-39deg.C), with rates of viral multiplication decreased at lower temperatures and increased at higher (McLean, 1975a,b). Therefore, although the A. aegypti is susceptible to infections from regions of the world in which the species is absent, the decreased temperature has such a negative effect on the EI period that this warm-climate vector would not be a competent vector in colder climates.
The infectivity threshold (IT), or concentration of virus that must be ingested by the vector in order for strains to become infective, varies among viruses and mosquito species. There are many barriers, such as salivary gland and mesenteronal barriers,[29] preventing infection of certain tissues, and so the most effective viruses have the lowest infectivity thresholds. However, a low IT is not necessarily a requirement for successful transmission if the virus produces very high levels of viremia in its vertebrate host, or if transovarial (vertical) transmission is possible (Hardy et al., 1983).
Transovarial transmission has been observed in many mosquito species and is considered to be a primary means by which some arboviruses are maintained during winter, dry seasons, and under adverse environmental conditions under which their arthropod hosts are either inactive or unable to survive. This is especially important in species in cooler climates and incapable of overwintering. Transovarial transmission was first observed for yellow fever virus in A. aegypti in 1905 (Rosen, 1981). Vertical transmission of SLE has also been widely demonstrated in eight species which occur in Florida. In this study, larval rearing temperatures affected such transmission of the virus in A. taeniorhynchus, with effective transmission at 18deg.C, but not at 27deg.C (Nayar et al., 1986). In a different study, vertical transmission of SLE by female A. albopictus and A. epactius occurred. Minimal infection rates of F1 adults reared at 18deg.C were much higher than in those reared at 27deg.C. This type of transmission is postulated to be a mechanism for overwintering of the SLE virus in temperate regions of North America. Dengue, IBE, and yellow fever viruses are also transmitted vertically at low levels by Aedes mosquitoes (Hardy et al., 1980).
It is difficult to make any generalizations regarding the geographic distributions of specific vectors without addressing each one individually. However, based on the affects of temperature changes, one can make a broad conclusion that species that are capable of overwintering or of transmitting vertically, and which have higher rates of transmission at lower EI temperatures, are more likely to be found in latitudes corresponding to temperate-zone to arctic-zone climates. The viruses for which they are specific will also be endemic to those areas.
DEFINITIVE HOST
The definitive host is the organism where the infectious agent grows and sexually multiplies. For these vector-borne diseases, the human host provides nutrition and the necessary environment for proliferation. The information on the five vector-borne diseases was reviewed; very little information was found specifically relating to the impact of climatic factors on modifying the role of the host in the infectious disease process.
...it is becoming increasingly evident that man rather than nature is likely to be responsible for the direct or indirect dispersal of an arbovirus over great distances. Although by no means proved, it is distinctly possible that recent major and singular epizootic epidemic outbreaks of Venezuelan equine encephalitis in Central and North America and of Rift Valley fever in Egypt were caused by human behaviour. In any event, it is now clear that most disease-producing arboviruses are not introduced into a known endemic region annually by migratory birds or other long-distance non-human travellers (WHO, 1985a).
Theoretically, there are two types of human activity that could impact the disease process and could in turn be modified by changes in climate: those activities that humans as society undertake and those activities that humans as individuals undertake.
Human activities in the societal sense, particularly as they relate to the environment, increase or decrease the amount of disease. Clearings, power lines, logging roads, and campsites frequently become heavily infested with ticks carrying RMSF (Hoogstraal, 1981). Increased use of irrigation may lead to increases in mosquito populations (Gratz, 1973).
The disappearance of plague from Uganda coincided with the introduction of antibiotics and DDT but probably had nothing to do with either, just as the virtual disappearance of malaria from the USA in the 1950s (which stood at 4 million cases per year in the 1930s) had little, if anything to do with the new anti-malarials and DDT. I well remember L.W. Hackett emphasizing this point when I was studying epidemiology at Berkeley in 1955. In the USA, as in Western Europe, changes in the ways of man were probably involved. Perhaps some of us should be making a special study of the factors involved in bringing about these great recessions with as much energy as we bring to the study of the factors responsible for the establishment of endemic and epidemic conditions (Gillett, 1985).
The environmental changes resulting from global climate change as well as the activities taken in response to these changes could potentially result in changes in the pattern of infectious disease. All the implications of global climate change are not yet known; however, within EPA's program to evaluate the potential impacts, consideration is being given to how hydrology, agriculture, forestry and infrastructure will change on a regional level (EPA, 1987). Potential changes which would result from sea level rise and changes in temperature and rainfall patterns include increased use of irrigation and impacts on wetlands and coastal, lake, and river ecosystems, such as geographic shifts and changes in composition; changes in the composition and growth rate of forests; and changes in crop yields, geographic distribution, and pesticide use. All of these changes have potential for impact on disease patterns and the growth and development of agent-vector pairs.
Perhaps of equal importance in the transmission of infectious disease in the United States is man's behavior as an individual. The best studied example of this is the use of air conditioning and television as it relates to mosquito-borne encephalitis in the San Joaquin Valley (Gahlinger et al., 1986). It was found that the use of these appliances decreased the amount of time people spent outdoors on summer evenings, the feeding time for the vector Culex tarsalis. The decreased prevalence of WEE and SLE in humans, together with the virus' continued presence in avian and equine populations, as well as the prevalence of the vector, can be accounted for only by the individual activities of man.
ADDITIONAL DISEASES
Participants at the workshop identified three additional diseases that they felt deserved attention in this report. A brief description of these diseases is given below.
Rift Valley fever (RVF) is a mosquito-borne, acute febrile disease of cattle, sheep, and man. It is characterized by fever, vomiting, hemorrhagic manifestations, vascular retinitis, encephalitis, and CNS complications (WHO, 1985b). RVF has long been recognized as a self-limited human febrile disease in veterinarians, butchers and shepherds. A 1975 RVF outbreak in South Africa produced the first known outbreak of RVF with encephalitis and fatal hemorrhagic fever (WHO, 1985a).
Although RVF has generally been confined to sub-Saharan Africa, outbreaks occurred in 1977-78 in the irrigated region of Egypt with devastating consequences. It is estimated that between 20,000 and 200,000 cases and 600 deaths occurred. Tests confirmed that RVF had not previously occurred in Egypt (WHO, 1985a).
RVF has not yet been reported outside Africa. However, the 1977-78 outbreak in Egypt demonstrated that it can occur in new areas with devastating results. Although the natural reservoir of RVF is unknown, it is believed that cattle and sheep act as amplifiers of the disease. At least 26 species of mosquito have been implicated as potential vectors of the virus by isolation in laboratory experiments including C. pipiens (WHO, 1985b). Its penetration into Egypt represents movement into a totally new ecosystem and combined with the severity of the human epidemic, it demonstrates disastrous potential for areas outside Africa.
Yellow fever, which shares clinical features with other hemorrhagic fevers, but has a more severe hepatic involvement, is a viral disease endemic to the tropical regions of the Americas and Africa. It is transmitted by mosquitoes from monkey to man and from man to man. At present, yellow fever has been reported from ten Latin American countries with most cases coming from Bolivia, Brazil, Columbia and Peru (WHO, 1985a; Monath, 1987).
Of particular importance is the outbreak of yellow fever in urban areas infested with A. aegypti, which is present in large parts of the United States. Several outbreaks of urban yellow fever have been traced to persons who were infected in forested areas and subsequently brought the disease to the city (WHO, 1985b).
Although an effective vaccine for yellow fever exists, supplies are not adequate. Should an outbreak occur in the continental United States, it has been predicted that supplies could not be produced in sufficient amounts to prevent an epidemic (Drs. Robert Shope and William Reeves, personal communication).
The changes in climate will increase the incidence and severity of skin infections and infestations. Skin infections, including dermatophytosis,[30] candidiasis,[31] streptococcal pyoderma, erythrasma, tinea versicolor,[32] and scabies are altered by temperature and humidity (Taplin et al., 1987). The importance of lifestyle and behavior upon contracting disease cannot, however, be dismissed.
Skin diseases were the single greatest cause of outpatient visits to Army medical facilities during the Vietnam War (Allen, 1977). When compared with temperature, relative humidity, and rainfall, outpatients' visits coincided with the mean monthly index values for rainfall and relative humidity, but were four months different from variations in monthly temperature (Figure 10). Thus, outpatient skin diseases were directly affected by rainfall and relative humidity, but not by temperature changes (Allen, 1977).
A major focus of the two-day workshop was to address the issue of what policies would need to be implemented to lessen the increase in morbidity and mortality from infectious disease caused by climate change. It was felt that there was already a significant amount of information known that could be preventively used to combat the spread of infectious disease in the United States.
The workshop participants felt that disease and vector surveillance programs are a cornerstone to dealing with the issues. Several states have in place surveillance programs which are adequate to deal with the problems (e.g., the California Encephalitis Surveillance Program). However, these programs are in danger of not being able to deal effectively with the situation because of a decrease in state and federal funding and a lack of trained personnel. There are also many areas in the country where these programs do not exist and may need to be established.
A major concern of the workshop was the deterioration in the centers of excellence that could be called upon to spearhead drives against the increases in infectious disease. For example, the number of arboviral research units in the United States has decreased, and it is the belief of the workshop attendees that the expertise may not be present in sufficient quantities to deal effectively with the situation.
The workshop attendees found that these issues are multidimensional in nature, and effective solutions would also have to be interdisciplinary. It was, therefore, felt that interdisciplinary teams need to be established so that the groups can be effectively integrated. The workshop felt that an effective integration necessitates working together for a period of time and, therefore, these groups need to be formed now.
The workshop participants expressed great concern that EPA's Endangered Species Act would limit pesticide use and thereby curtail the ability of various agencies to control vector-borne disease.
As has been discussed, climate has a great impact on patterns of infectious disease in the extent of their geographic radius as well as in their seasonal duration. Although the importance of certain climatic conditions has been defined for a number of infectious diseases, it is not well understood how changes in climate, particularly gradual change, will affect disease patterns.
This report has reviewed what is known on many of the issues and has presented our best estimates of how climatic changes could alter the patterns of infectious disease in the United States. Without knowing exactly what changes in climate will occur (information which the EPA is in the process of generating), it is not possible to predict what the impacts of the global climate changes due to the emissions of "greenhouse gases" will be. In addition, recommendations are given below with regard to the data gaps which need to be filled.
ADDITIONAL RESEARCH NEEDED
In the course of developing this report, it was realized that there are many areas that need additional research. These research areas are outside the traditional health literature, but are issues that directly impact on the question of climatic change and infectious disease.
Footnotes
13 A. quadrimaculatus is highly susceptible to both P. vivax and P. falciparum.
14 A. albopictus has been shown experimentally to transmit dengue transovarially.
18 WEE has an alternative cycle in jackrabbits and A. melanimon.
24 "Dangerous contagious diseases" include AIDS, tuberculosis, VD, and leprosy.
27 The same study indicated a correlation of lower body fat with more normal ovarian development.
GLOSSARY
anopheline mosquitoes - mosquitoes of the genus Anopheles; many are malarial vectors.
antibiotic - a chemical derived from a fungus or bacteria which is inhibitory to other microorganisms.
antigen - substance which induces an immune response upon contact with the immune system.
arbovirus - arthropod-borne virus.
arthropod - member of the invertebrate phylum Arthropoda which includes insects, crustaceans, spiders, and ticks.
candidiasis - infection caused by yeast species of the genus Candida, often Candida albicans (Stein, 1983).
CDC - Centers for Disease Control.
CE - California encephalitis. chloroquinine - see quinine.
CNS - central nervous system.
coagulation - the clotting of blood.
coccobacillus - a type of bacteria
definitive host - organism in which the infectious agent grows and sexually multiplies; for the diseases discussed in this paper, the definitive host is often a human.
Dengue fever - infectious disease caused by the dengue virus, which is transmitted by mosquitoes.
DEN-1, -2, -3, -4 - the four serotypes of the dengue virus.
dermatophytosis - infection caused by fungus of one of three genera: Microsporum, Trichophyton, or Epidermophyton; may be referred to as ringworm or Tinea (Stein, 1983).
desiccation- drying.
DHF - dengue hemorrhagic fever.
diapause - physiological state of suspended activity or arrested development that facilitates survival through a period of unfavorable conditions, but is initiated before the onset of these conditions (Bailey, 1982).
dysarthria - malformation or disturbance of a joint due to emotional stress, paralysis, or spasticity of muscles.
edema - swelling as a result of accumulation of fluid.
EEE - eastern equine encephalitis.
encephalitis - inflammation of the brain.
endemic - present in a certain area or among a specific population.
epithelium - layer of cells covering all free surfaces of an organism.
epizootic - a disease which simultaneously attacks or is present in a large number of animals.
erythema chronicum migran (ECM) - skin lesion consisting of an inflamed, red ring with advancing hardened edges leaving a central clear area; emanates from an insect bite; characteristic of Lyme disease.
erythrasma - eruption of reddish brown patches as a result of Corynebacterium minutissimum.
etiology - cause of disease.
extrinsic incubation (EI) period - interval between host's ingestion of the virus and subsequent transmission through biting; varies for each virus and vector.
febrile - relating to fever.
gram-negative - a defining characteristic of a group of bacteria; after being Gram stained, gram-negative cells do not readily retain crystal violet dye.
helminth - an intestinal parasite.
hemorrhagic - relating to or characterized by bleeding.
hepatic - relating to the liver.
host - organism from which infectious agent gains sustenance.
indigenous - originating in a particular area.
infectious feeding - introduction of the agent to the vector when the vector is feeding on the blood of an infected organism.
infectious agent - the virus, bacteria, protozoan, or other microorganism which induces disease.
infectivity threshold - concentration of virus that must be ingested by the vector in order for strains to become infective; varies among viruses and vectors.
JBE - Japanese B encephalitis.
larva - immature form of certain organisms (e.g., insects and ticks) which emerge from the egg.
Lyme disease - inflammatory infectious disease caused by the tick-transmitted spirochete, Borrelia burgdorferi.
meningeal - relating to the membranes surrounding the spinal cord and brain.
meningitis - inflammation of the membranes of the brain and/or spinal cord.
mesenteronal - relating to the layers of tissue surrounding the abdominal viscera.
mesic - characterized by a moderate amount of moisture.
nymph - immature form of certain organisms (e.g., ticks); the larva molts to become a nymph, which is adult like in form though usually smaller.
pantropical - relating to areas which are predominantly tropical.
parenchymal - relating to the distinguishing cells of a gland or organ.
paresis - partial or incomplete paralysis.
periorbital - relating to the area around the eye socket.
photoperiod - duration of light.
plasmodia - members of the family Plasmodiae; blood parasites of vertebrates.
protozoa - unicellular eukaryotic microorganisms.
pupa - inactive form of an organism during which the larva transforms into an adult.
questing period - time when ticks--either nymphs or adults--are seeking hosts.
quinine/chloroquinine - antimalarial chemical effective against the asexual and red blood cell-attacking forms of the plasmodia.
resistance - ability of an organism to remain unaffected by a toxic substance.
retinitis - inflammation of the retina.
rickettsia - a type of coccobacillus (bacteria); all but one of this group are vector-borne parasites.
RMSF (Rocky Mountain spotted fever; also know as tick typhus) - infectious disease caused by the tick-borne bacteria Rickettsiae rickettsii.
RVF - Rift Valley fever.
scabies - skin irritation accompanied by intense itching caused by the female Sarcoptes scabiei var. hominis burrowing into the skin.
sequela - effect of a disease.
serotype - subdivision of a species or virus identifiable on the basis of antigenic character.
SLE - St. Louis encephalitis.
spirochete - spiralling bacteria streptococcal pyoderma - a pus-forming skin infection caused by a member of the genus Streptococcus.
systemic - relating to the organism as a whole.
tinea versicolor - infection of the superficial layer of the epidermis by the yeast Melassezia furfur (Stein, 1983).
transovarial transmission - passage of infectious agent to eggs within the ovaries; larvae are subsequently infected.
transstadial transmission - transmission of agent between different stages in the life history of an organism.
vector - organism which transmits the infectious agent to the host; examples include ticks and mosquitoes.
VEE - Venezuelan equine encephalitis.
venereally - relating to sexual intercourse.
vertical transmission- see transovarial transmission.
viremia - the presence of a virus in the bloodstream.
WEE - western equine encephalitis.
yellow fever - acute, harmful infectious disease caused by a mosquito-borne virus.
REFERENCES
Allen, A.M. Skin Diseases in Vietnam. 1965-72. United States Army, 1977.
American Public Health Association. APHA. Control of Communicable Diseases in Man (ed.) Berenson, A.S. (1980)
Anon. Leads from the MMWR, Imported and indigenous Dengue fever - United States, 1986. JAMA 258:1712-1713 (1987a).
Anon. Leads from the MMWR; Dengue in the Americas, 1985. JAMA 257:166 (1987b).
Arfaa, F. Intestinal parasites among Indochinese refugees and Mexican immigrants resettled in Contra Costa County, California J Fam Prac 12:223-226 (1981).
Bailey, C.L., Faran, M.E., Gargan, T.P., and Hayes, D.E. Winter survival of blood-fed and nonblood-fed Culex pipiens L. Am J Trop Med Hyg 31:1054-1061 (1982).
Barry, M., Craft, J., Coleman, D., Coulter, H.O., and Horwitz, N. Clinical findings in Southeast Asian refugees. JAMA 249:3200-3203 (1983).
Bos, E. Estimate of the number of illegal aliens: An analysis of the sources of disagreement. Pop Res Pol Rev 3:239-254 (1984).
Botkin, D.B., Janak, F.F. and Wallis, J.R. Some ecological consequences of a computer model of forest growth. J Ecol 60:849-872 (1972).
Bowen, G S., Schultz, T.L., Hayne, C., and Parkin, W.E. Focus of Lyme disease in Monmouth County, New Jersey. Am J Epi 120:387-394 (1984).
Bozeman, M.F., Shirai, A., Humphries, J.W., et al. Ecology of Rocky Mountain spotted fever. 2. Natural infection of wild mammals and birds in Virginia and Maryland. Am J Trop Med Hyg 16:48 (1967).
Braunwald, E., Isselbacher, K.I., Persdorf, R.G., Wilson, J.D., Martin, J.B., and Fauci, A.S. Harrison's Principles of Internal Medicine. Eleventh Edition. McGraw-Hill (1988).
Brunetti, R., Fritz, R.F., and Hollister, A.C. An outbreak of malaria in California, 1952-1953. Am J Trop Med 3:779-788 (1954).
Burgdorfer, W., Lane, R.S., Barbour, A.G., Gresbrink, R A., and Anderson, J.R. The western black-legged tick, Ixodes pacificus: A vector of borrelia burgdorferi. Am J Trop Med Hyg 34:925-930 (1985).
Cantanzaro, A. and Moser, RJ. Health status of refugees from Vietnam, Laos and Cambodia. JAMA 247:1303-1308 (1982).
CDC. Health Status of Indochinese refugees. MMWR 28:385-398 (1979a).
CDC. Survey of intestinal parasites-Illinois. MMWR 28:346 (1979b).
CDC. Viral hepatitis type B, tuberculosis and dental care of Indonesian refugees. MMWR 29:1-3 (1980c).
CDC. Health screening resettled Indochinese refugees-Washington, D.C., Utah. MMWR 29:4-11 (1980e).
CDC. Dengue-Cuba MMWR 30:317 (1981).
CDC. USPHS Guidelines for medical examination of aliens. June, 1984.
CDC. Plasmodium viral malaria-San Diego County, California, 1986. MMWR 35:679-681 (1986a)
CDC. Update: Aedes albopictus infestation-United States. MMWR 35:649-651 (1986b).
CDC. Aedes albopictus introduction-Texas. MMWR 35:141-142 (1986c).
CDC. Aedes albopictus infestation-United States. MMWR 35:493-495 (1986d).
CDC. Dengue - the Americas, 1984. MMWR 35:51-57 (1986e).
CDC. Annual Summary, 1984. MMWR 33:54 (1986f).
CDC. Update: Aedes albopictus infestation-United States. MMWR 36:769-773 (1987).
Corwin, A.F. The numbers game: Estimates of illegal aliens in the United States, 1970-1981. Prepared for the House Judiciary Committee. Feb 1982.
D'Angelo, L.J., Bregman, D.J. and Winkler, W.G. Rocky Mountain spotted fever in the United States: Use of age-specific incidence to determine public health policy for a vector-borne disease. So Med J 75:3 (1982)
Dauer, C.C. and Faust, E.C. Symposium on malaria - Part 2. Malaria mortality in the United States, with especial reference to the Southeastern United States. So Med J 30:939-943 (1937).
de Zulueta, J. Man and malaria In: Stanley, N.F., and Joske, R A. (eds), Changing Disease Patterns and Human Behavior, Academic Press, New York (1980).
Ehrenkranz, N.J., Ventura, A.K., Cuadrado, R.R., Pond, W.C., and Porter, J.E. Pandemic dengue in Caribbean countries and the southern United States-past, present, and potential problems. N Eng J Med 285:1460-1469 (1971).
Eldridge, B.F. The effect of temperature and photoperiod on blood-feeding and ovarian development in mosquitoes of the Culex pipiens complex. Am J Trop Med Hyg 17:133-143 (1968).
EPA/UNEP. Effects of changes in stratospheric ozone and global climate. Vol. 1-4 (1986).
EPA. U.S. Environmental Protection Agency. Work Plan for a Report to Congress on the Effects of Global Climate Change. September 16, 1987 (1987).
Farhang, A.A., and Traub, R. Transmission of murine typhus rickettsiae by Xenopsylla cheopsis, with notes on experimental infection and effects of temperature. Am J Trop Med Hyg 34:555-563 (1985).
Faust, E.C. The 1930 status of malaria in the southern United States, as determined by mortality data. So Med J 25:544-548 (1932).
Faust, E.C. Malaria mortality in the southern United States for the year 1936. So Med J 31:816-818 (1938).
Faust, E.C. Malaria mortality in the southern United States for the year 1937. Am J Trop Med 19:447-455 (1939).
Finley, K. and Longshore, W A. A progress report on five years of follow up of western equine and St. Louis encephalomyelitis cases in California with special reference to children. Trans Amer Neur Soc 83:45-49 (1958).
Gahlinger, P.M., Reeves, W.C., and Milby, M.M. Air conditioning and television as protective factors in arboviral encephalitis risk. Am J Trop Med Hyg 35:601-610 (1986).
Garvie, M.B., McKiel, J.A., Sonenshine, D.E., and Campbell, A. Seasonal dynamics of American dog tick, Dermacentor variabilis (Say), populations in southwestern Nova Scotia. Can J Zool 56:28-39 (1978).
Gillett, J.D. Direct and indirect influences of temperature on the transmission of parasites from insects to man. In: Taylor, A.E.R. and Muller, R. (eds), The Effects of Meteorological Factors upon Parasites, Blackwell Scientific Publications, London (1974).
Gillett, J.D. The behaviour of Homo sapiens, the forgotten factor in the transmission of tropical disease. Trans R Soc Trop Med Hyg 79:12-20 (1985).
Gratz, N.G. Mosquito-borne disease problems in the urbanization of tropical countries. CRC Crit Rev Environ Control 3:455-495 (1973).
Gubler, D.J. Dengue in the United States. MMWR 33(1SS):9SS-12SS (1983).
Habicht, G.S., Beck, G., and Benach, J.L. Lyme disease. Sci Am 256:78-83 (1987)
Hardy, J.L., Rosen, L., Kramer, L.D., Presser, S.B., Shroyer, D A. and Turell, M.J. Effect of rearing temperature on transovarial transmission of St. Louis encephalitis virus in mosquitoes. Am J Trop Med Hyg 29:963-968 (1980).
Hardy, J.L., Houk, EJ., Kramer, L.D., and Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Ann Rev Entomol 28:229-62 (1983).
Hess, A.D., Cherubin, C.E., and LaMotte, L.C. Relation of temperature to activity of Western and St. Louis encephalitis viruses. Am J Trop Med Hyg 12:657-667 (1963).
Hoogstraal, H. Changing patterns of tick-borne diseases in modern society. Ann Rev Entomol 26:75-99 (1981).
Hoogstraal, H., Kaiser, M.N., Traylor, M.A., Gaindy, E., and Gaber, S. Ticks (Ixodidae) on birds migrating from Europe and Asia to Africa, 1959-1961. Bull WHO 28:235-262 (1963).
Immigration and Naturalization Service. INS. Statistical Yearbook (1985).
Johnson, R.T. Viral Infections of the Nervous System, Raven Press, New York, pp. 107-119 (1977).
Jones, M.J., Thompson, J.H., and Brewer, N.S. Infectious diseases of Indochinese refugees. Mayo Clin Proc 55:482-488 (1980)
Kramer, L.D., Hardy, J.L. and Presser, S.B. Effect of temperature of extrinsic incubation on the vector competence of Culex tarsalis for western equine encephalomyelitis virus. Am J Trop Med Hyg 32:1130-1139 (1983).
Lane, R S. and Burgdorfer, W. Potential role of native and exotic deer and their associated ticks (Acari: Ixodidae) in the ecology of Lyme disease in California, USA. Zbl Bakt Hyg A 263:55-64 (1986).
Longstreth, J.D., Lill, P., Wiseman, JA., and Laurenson, J. UV-B Induced Immunosuppression: Potential Impacts on Infectious Disease. Report prepared for U.S. EPA (1987).
Macdonald, G. The Epidemiology and Control of Malaria. Oxford University Press, Oxford (1957).
Main, A.J., Carey, A.B., Carey, M.G., and Goodwin, R.H. Immature Ixodes dammini (Acari: Ixodidae) on small animals in Connecticut, USA. J. Med Entomol 19:655-664 (1982).
Mauser, J.S., and Kramer, S. Epidemiology: An Introductory Text. (2nd ed.) W.B. Saunders Co., St. Louis, MO (1985).
McDade, J.E., and Newhouse, V.F. Natural history of Rickettsia rickettsii. Ann Rev Microbiol 40:287-309 (1986).
McEnroe, W.D. The restriction of the species range of Ixodes scapularis, Say, in Massachusetts by fall and winter temperature. Acarologia 18:618-625 (1977).
McGlynn, K.A., Lustbader, E.D., and London, W.T. Immune responses to hepatitis B virus and tuberculosis infections in Southeast Asian refugees. Am J Epi 122:1032-1036 (1985).
McLean, D.M., Grass, P.N., Judd, B.D, Cmirlova, D., and Stuart, K.M. Natural foci of California encephalitis virus activity in the Yukon Territory. Can J Pub Health 68:69-73 (1977).
McLean, D.M., Miller, M.A., and Grass, P.N. Dengue virus transmission by mosquitoes incubated at low temperatures. Mosq News 35:322-327 (1975a).
McLean, D.M., Grass, P.N., Miller, M A., and Wong, K.S.K. Arbovirus growth in Aedes aegypti mosquitoes throughout their viable temperature range. Arch Virol 49:49-57 (1975b).
Monath, T.P. Yellow fever: A medically neglected disease. Report on a seminar. Rev Infect Dis 9:165-175 (1987).
Monath, T.P. Epidemiology. In: Monath, T.P. (ed.), St. Louis Encephalitis, APHA, Washington, D.C. pp. 239-312 (1980).
National Academy of Sciences (NAS). Changing climate. National Academy Press, Washington, D.C. (1983).
Nayar, J.K., Rosen, L., and Knight, J. Experimental vertical transmission of Saint Louis encephalitis virus by Florida mosquitoes. Am J Trop Med Hyg 35:1296-1301 (1986).
Nicholls, N. A method for predicting Murray Valley encephalitis in Southeast Australia using the southern oscillation. Aust J Exp Biol Med Sci 64:587-594 (1986).
Peters, R.L., and Darling, J.D.S. Potential effects of greenhouse warming on natural communities. In: Titus, J. (ed.), Effects of Changes in Stratospheric Ozone and Global Climate. Vol. 3, EPA, Washington, D.C. pp. 137159 (1986).
Rosen, L. Transmission transovarienne des arbovirus par les moustiques. Med Trop 41:23-29 (1981).
Schulze, T.L., Bowen, G.S., Lakat, M.F., Parkin, W.E., and Shisler, J.K. Seasonal abundance and hosts of Ixodes dammini (Acari: Ixodidae) and other Ixodid ticks from an endemic Lyme disease focus in New Jersey, USA. J Med Entomol 23:105-109 (1986).
Schulze, T.L., Bowen, G.S., Bosler, E.M., Lakat, M.F., Parkin, W.E., Altman, R., Ormiston, B.G., and Shisler, J.K. Amblyomma americanum: A potential vector of Lyme disease in New Jersey. Science 224:601-603 (1984).
Shope, R.E. Arbovirus-related encephalitis. Yale J Biol Med 53:93-99 (1980).
Solomon, A.M. and West, D.C. Simulating forest ecosystem responses to expected climate change in eastern North America. Applications to decision making in the forest industry. Publ. No. 2398, Environmental Science Division, Oak Ridge National Lab, Oak Ridge, Tenn. (1984).
Sonenshine, D.E., Peters, A.H., and Levy, G.F. Rocky Mountain spotted fever in relation to vegetation in the Eastern United States. Am J Epi 96:59-69 (1972).
Sonenshine, D.E., and Clifford, C.M. Contrasting incidence of Rocky Mountain spotted fever in ticks infesting wild birds in eastern U.S. Piedmont and coastal areas, with notes on the ecology of these ticks. J Med Entomol 10:497-502 (1973).
Spence, L., Artsob, H., Grant, L., and Th'ng, C. St. Louis encephalitis in southern Ontario: Laboratory studies for arboviruses. Can Med Assoc J 116:35-37 (1977).
Spielman, A., Wilson, M.L., Levine, J.F., and Piesman, J. Ecology of Ixodes dammini-borne human babesiosis and Lyme disease. Ann Rev Entomol 30:439-460 (1985).
Spielman, A., Levine, J.F., and Wilson, M.L. Vectorial capacity of North American Ixodes ticks. Yale J Biol Med 57:507-513 (1984).
Steere, A.C., Burtenhagen, N.H., Craft, J.E., et al. The early clinical manifestations of Lyme disease. Ann Intern Med 99:76-82 (1983a).
Steere, A.C., and Malawista, S.E. The epidemiology of Lyme disease, In: Lawrence, N.E., and Schulman, L.E. (eds). Current Topics in Rheumatology: Epidemiology of the Rheumatic Diseases, Gower Med Publ Ltd., New York p 33 (1983b).
Steere, A.C., and Malawista, S.E. Cause of Lyme disease in the United States: Locations correlated with distribution of Ixodes dammini. Ann Intern Med 91:730-733 (1979).
Stein, D.H. Superficial fungal infections. Ped Clin N A 30:545-561 (1983).
Taplin, D., Meinking, T.C., and Mertz, P.M. Skin infections in tropical climates: Current patterns and future predictions. Manuscript in preparation. (1987).
Thompson, W.H. and Beaty, B.J. Venereal transmission of La Crosse virus from male to female Aedes triseriatus. Am J Trop Med Hyg 27:187-196 (1978).
Turell, M.J., Reeves, W.C. and Hardy, J.L. Evaluation of the efficiency of transmission of California encephalitis viral strains in Aedes dorsalis and Aedes melanimon. Am J Trop Med Hyg 31:382-388 (1982).
Usinger, R.L. Entomological phases of the recent dengue epidemic in Honolulu. Pub Health Rep 59:423-430 (1944).
Watts, D.M., Burke, D.S., Harrison, B.A., Whitmire, R.E., and Nisalak, A. Effect of temperature on the vector efficiency of Aedes aegypti for Dengue 2 virus. Am J Trop Med Hyg 36:143-152 (1987)
Westrom, D.R., Lane, R.S., and Anderson, J.R. Ixodes pacificus (Acari: Ixodidae): Population dynamics and distribution on Columbian black-tailed deer (Odocoileus hemionus Columbianus). J Med Entomol 22:507-511 (1985).
WHO. Arthropod-borne and rodent-borne viral diseases. Tech Rep Ser #719. World Health Organization Geneva, Switzerland (1985a).
WHO. Viral hemorrhagic fevers. Tech Rep Ser #721. World Health Organization, Geneva, Switzerland (1985b).
WHO. The biology of malaria parasites. Tech Rep Ser #743. World Health Organization, Geneva, Switzerland (1987).
Wilkinson, P.R. The distribution of Dermacentor ticks in Canada in relation to bioclimatic zones. Can J Zool 45:517-537 (1967).
Williams, L.L. Report of the subcommittee on malaria prevention activities, 1937. So Med J 31:818-819 (1938).
Williams, L.L. Symposium on malaria - Part 2; malaria prevention activities, 1936. So Med J 30:938-939 (1937).
Yasuno, M., and Tonn, RJ. A study of biting habits of Aedes aegypti in Bangkok, Thailand. Bull WHO 43:319-325 (1972).
GOAL:
The primary goals of the workshop were to determine which infectious diseases would become significantly greater human health problems due to climate change and to determine what public health policy decisions can presently be made to lessen the morbidity and mortality of these infectious diseases due to changes in climate. The workshop also had the goal the defining of a research agenda for the future.
ISSUES:
1. Which infectious diseases pose the greatest threat in terms of morbidity and mortality within the United States due to climate change?
2. To what climate factors do these infectious diseases show the greatest sensitivity and which elements (i.e., agent, vector, reservoir host, human host) are most sensitive?
3. How significant could the infectious disease problem become in the future if temperatures rise by 2 to 4deg.C and precipitation changes by +/- 20 percent? What would a 1deg.C rise and a 10% change in precipitation do?
4. Are there any short or long term public health policy decisions currently being made which are likely to be impacted by climate change? Examples might be implementation of health screening of immigrants, or planning and/or implementing vector control programs.
5. What research needs are most critical in order to develop an assessment of the problem?
6. How would your answers change if these questions were applied to other developed countries? To underdeveloped countries?
The consensus of the workshop was that the United States would again have endemic (and epidemic) malaria and dengue (which is already present in Puerto Rico) in significantly greater numbers than is true today. These diseases would be mainly focused in the south-southeast and California, although the workshop did not rule out that these diseases would also occur in other areas of the United States.
The workshop also believed that yellow fever would again become a problem, even though a vaccine exists for the disease. This is so because very little of the vaccine exists in the United States (or throughout the world) and the supplies would easily be exhausted in the United States should a major outbreak occur.
It was generally felt that tick-borne diseases such as Lyme disease and RMSF would not become a greater problem due to climate change, but that the geographic focus of the diseases could change.
The workshop felt that temperature was the most important climatic variable and that rainfall was the second most important variable. Small changes in these variables significantly impact on the geographic range and temporal duration of the vector which was considered to be the element most sensitive to climatic change.
The workshop felt that these relatively small changes would have a dramatic impact in changing patterns of infectious disease. This is due to the real impact that small changes in climate can have on the vector. It was felt that the most important impact would be that small increases in temperature would increase the geographic range in which the vector would be present throughout the entire year.
Several public health decisions are presently being made which will undermine the ability of the United States to adequately deal with the changes in patterns of infectious disease brought about by climatic change.
One element is the lessening of the disease surveillance system in the United States. Without good surveillance data, preventive measures cannot be put into operation effectively.
There are becoming less centers of excellence, such as the arboviral research centers, in the United States that are conducting research on these questions and that will be able to advise and respond as necessary.
Due to the lack of career paths and adequate funding, fewer and fewer individuals, both in the United States and throughout the world, are being trained in vector eradication techniques. As climate change increases the importance of these programs, there will be a lack of adequately trained personnel to deal with the problem.
The issues discussed are multi-dimensional in nature. There are at present no multi-dimensional centers or groups that have integrated epidemiologists, entomologists, behavioral scientists and physicians that will be able to respond to the new problems raised by climate change.
The workshop felt that the most critical research needs were the collection of better data as to the incidence of infectious disease, particularly in those groups which traditionally do not use the health care system, i.e., the poor and recent immigrants.
It was also felt that greater surveillance of the various disease-carrying vectors is needed so that abatement programs can be maintained or developed as needed.
In relation to other developed countries, it was generally felt that the same or similar problems as predicted for the United States would occur in those countries. Similar influxes of immigrants from highly endemic areas occur throughout Europe as in the United States, and a similar strain on the public health systems of those countries is presently taking place.
The workshop believed that the impact of climate change on patterns of infectious disease in underdeveloped countries would be significantly greater than in developed countries. This was because the public health systems and vector abatement programs in these countries are already overburdened, and any additional burdens on these systems will result in significantly greater incidence of disease. In addition, the sanitation and nutrition levels in many of the underdeveloped countries will promote a greater incidence of infectious disease than in the more developed countries.
Dr. Joan AronDepartment of Population Dynamics Johns Hopkins University School of Public Health 615 North Wolfe Baltimore, MD 21205 301/955-3708
Dr. Thomas Chambers St. Jude's Children's Medical Research Center Department of Virology Box 318 Memphis, TN 38101 901/522-0300
Dr. James Childs Department of Immunology and Infectious Disease Johns Hopkins University School of Public Health 615 North Wolfe Baltimore, MD 21205 301/955-3708
Dr. Jeanne Courval Division of Epidemiology School of Public Health Columbia University 600 West 168 Street New York, NY 10032 212/305-3921
Ms. Eleanor Cross Naval Medical Research Mail Stop 7 Bethesda, MD 20814 301/295-1731
Ms. Margo Daniel United States Environmental Protection Agency Washington, DC 202/382-2776
Dr. I.F. Goldstein School of Public Health Columbia University 600 West 168 Street New York, NY 10032 212/928-7674
Dr. John Grayzell ST/RD/RRD Room 608 Dept. of State Washington, DC 20523 202/235-8860
Dr. Dan Haile USDA 1600 SW 23rd Drive Box 14565 Gainesville, FL 32604 904/374-5928
Dr. Charles Hughes Department of Family and Preventive Medicine University of Utah Medical Center Salt Lake City, UT 84132 801/581-5310
Ms. Terri Lavin Department of Geography University of Delaware Newark, DE 19716
Dr. Janice Longstreth ICF/Clement Associates 9300 Lee Highway Fairfax, VA 22031 703/934-3102
Ms. Terry Meinking Department of Dermatology and Cutaneous Surgery University of Miami School of Medicine Box 016960 R-117 Miami, FL 33101 305/547-6214
Dr. Greg Mertz Tufts School of Veterinary Medicine 200 Westboro Road North Grafton, MA 10536 617/839-5302
Ms. Marilyn Milby School of Public Health Warren Hall University of California Berkeley, CA 94720 415/642-3938
Dr. William Reeves School of Public Health Warren Hall University of California Berkeley, CA 94720 415/642-3933
Dr. Paul Reiter CDC Laboratory GPO Box 4532 San Juan, PR 00936 809/781-3636
Dr. Ronald Schwarz TRAIN, Inc. 2816 Cheswolde Baltimore, MD 21209 301/358-9561
Dr. Robert Shope Yale University School of Medicine Department of Epidemiology and Public Health Box 3333 New Haven, CT 06510 203/785-4821
Dr. David Slade Deputy Director Physical and Technical Research Div. ER-74 Office of Health and Environmental Research Department of Energy Washington, DC 20545 202/353-4375
Dr. Chip Stem Tufts School of Veterinary Medicine 200 Westboro Road North Grafton, MA 10536 617/839-5302
Dr. David Taplin Department of Dermatology and Cutaneous Surgery University of Miami School of Medicine Box 016960 R-117 Miami, FL 33101 305/547-6214
Ms. Kathleen Valimont Department of Geography University of Delaware Newark, DE 19716
Dr. Byron Wood TGS Technology Inc. Mail Stop 242-4 NASA Ames Research Center Moffit Field, CA 94035 415/694-6184
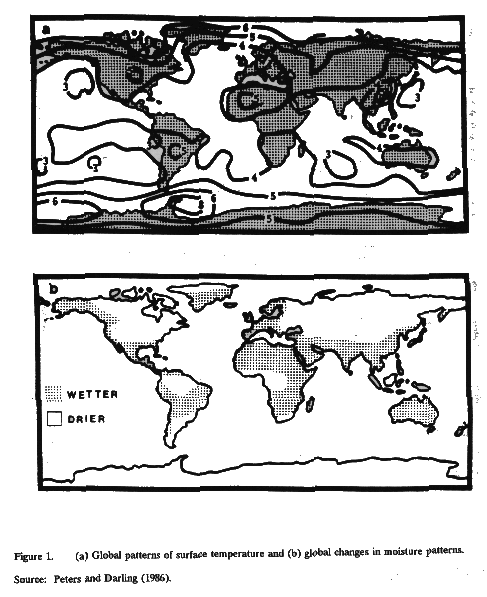{kind=link}
{kind=link}
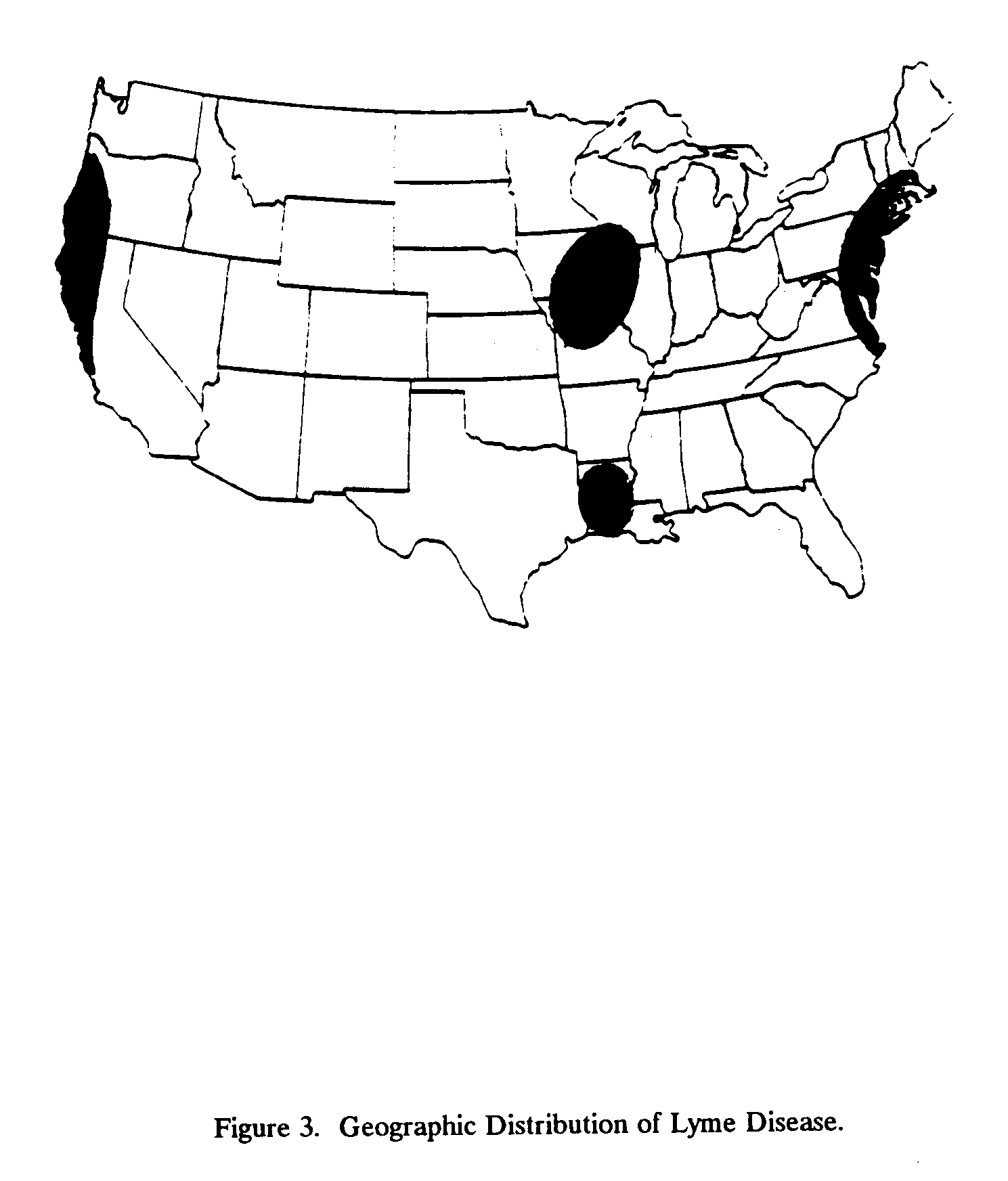{kind=link}
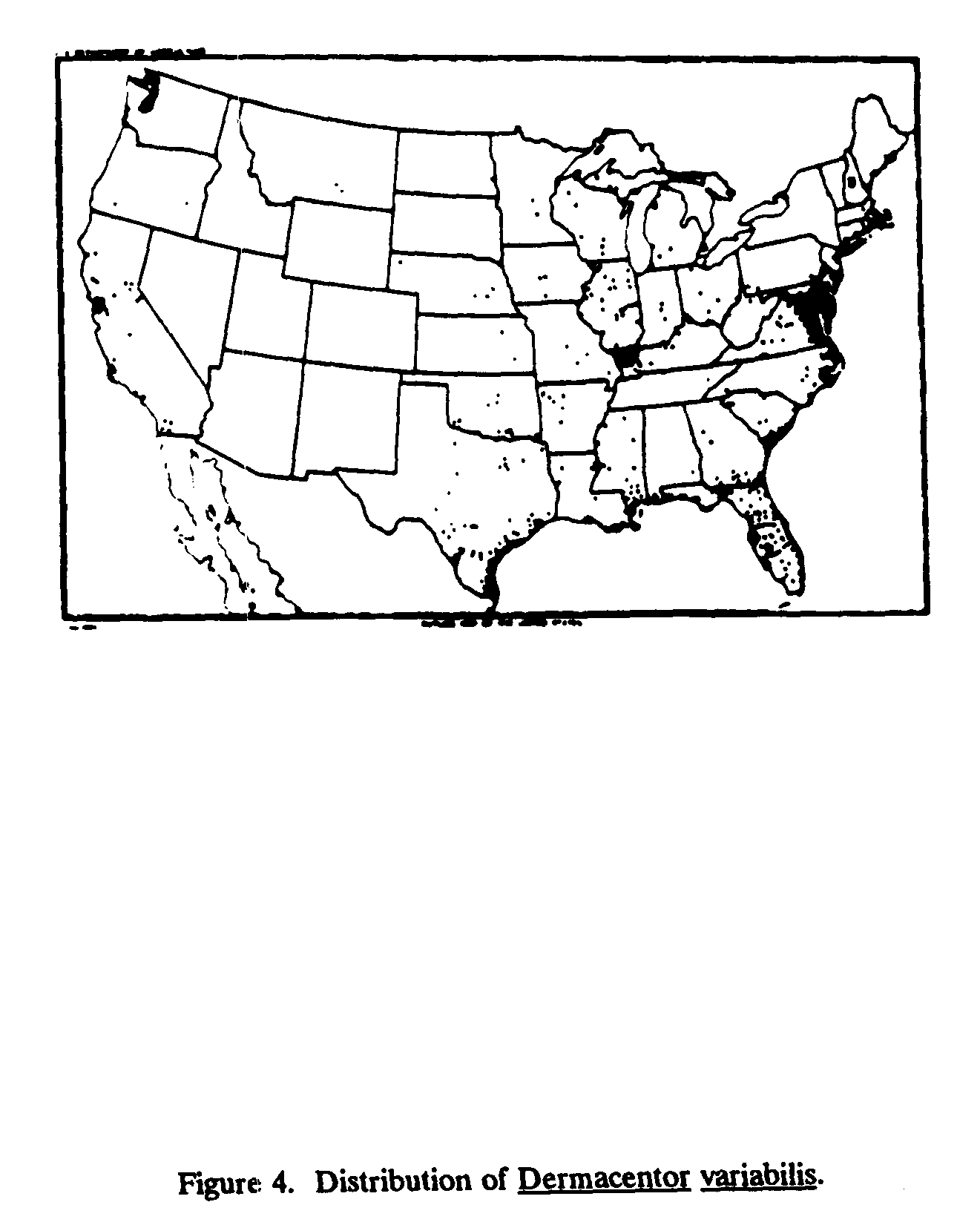{kind=link}
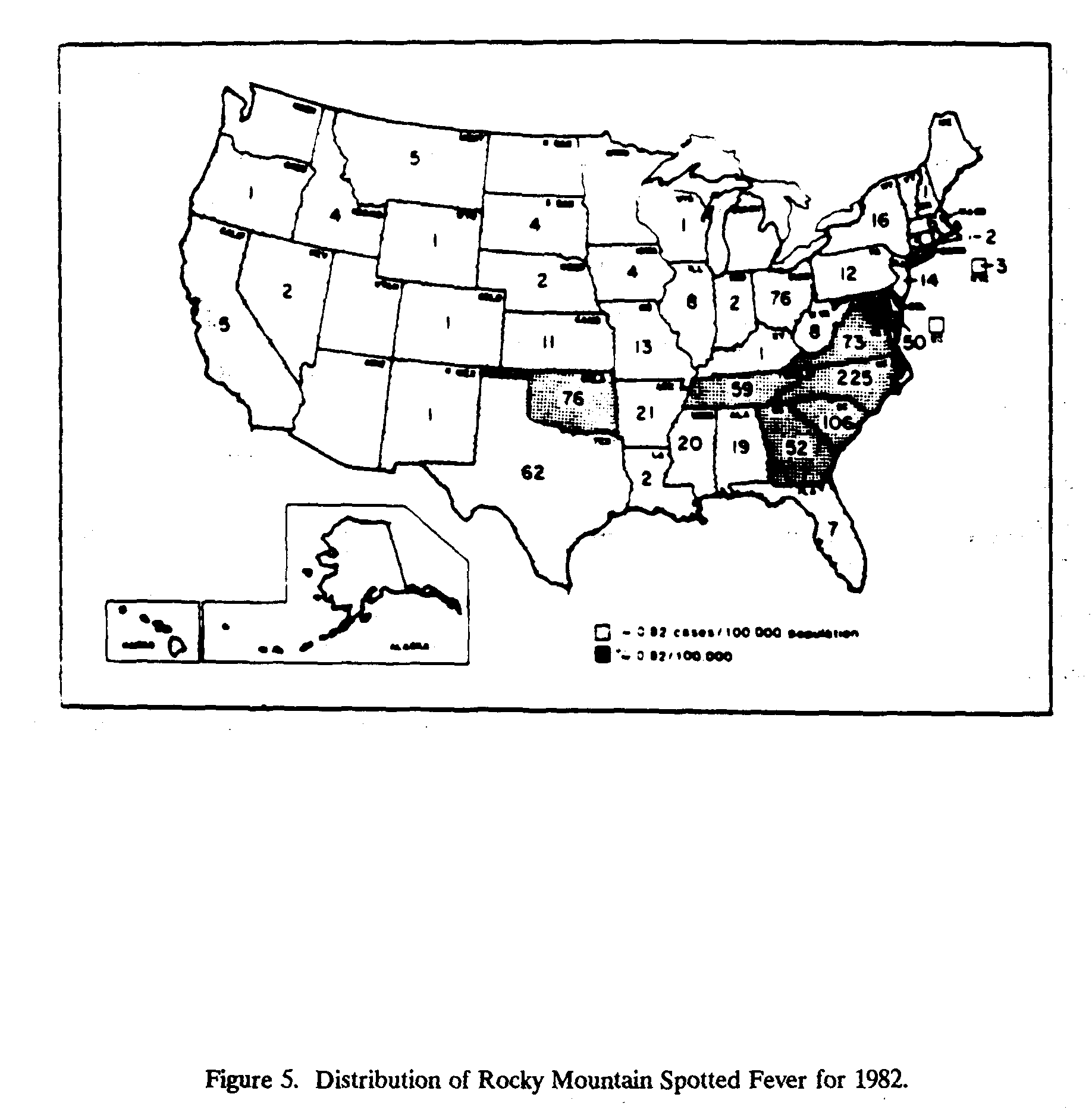{kind=link}
{kind=link}
{kind=link}
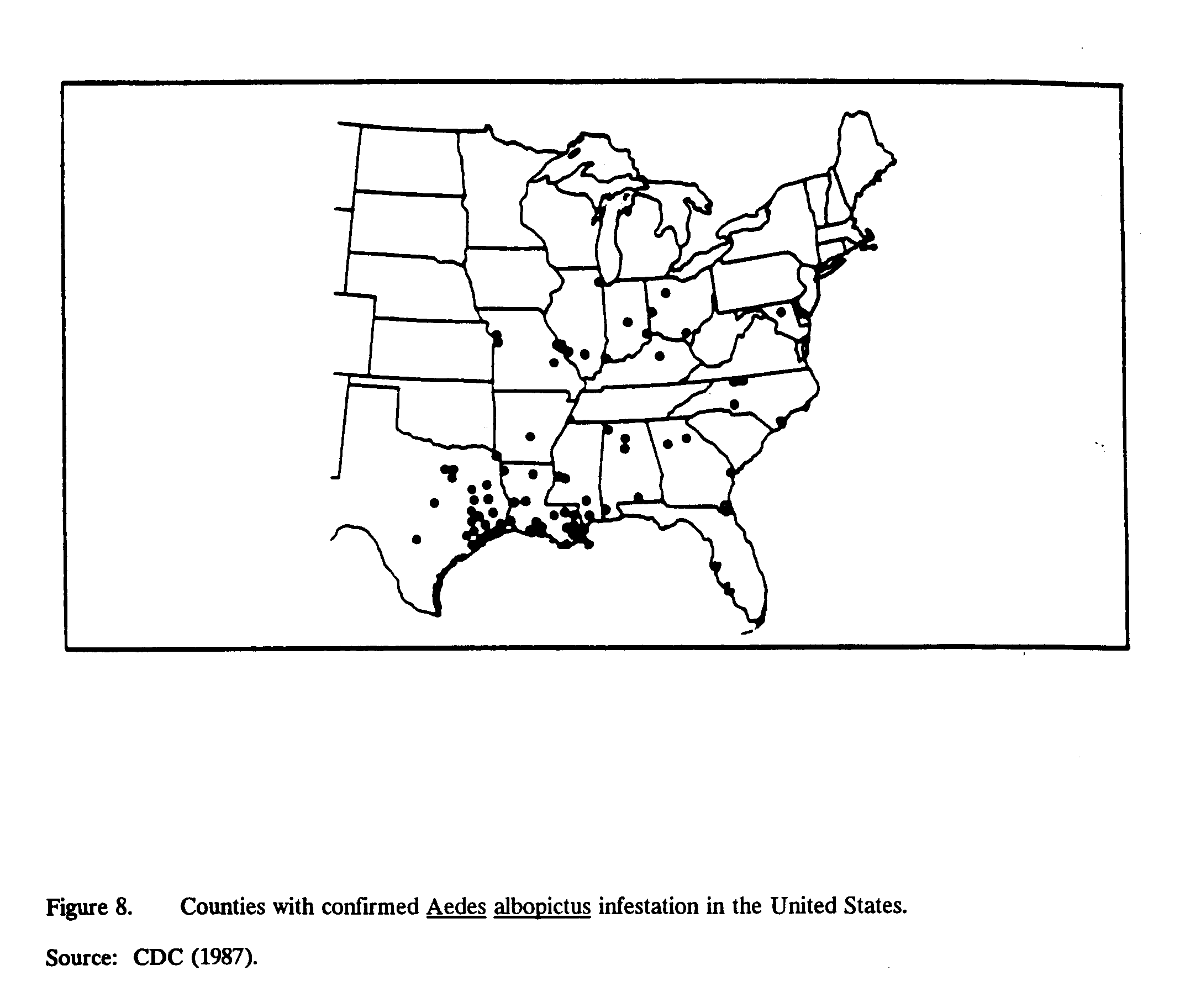{kind=link}
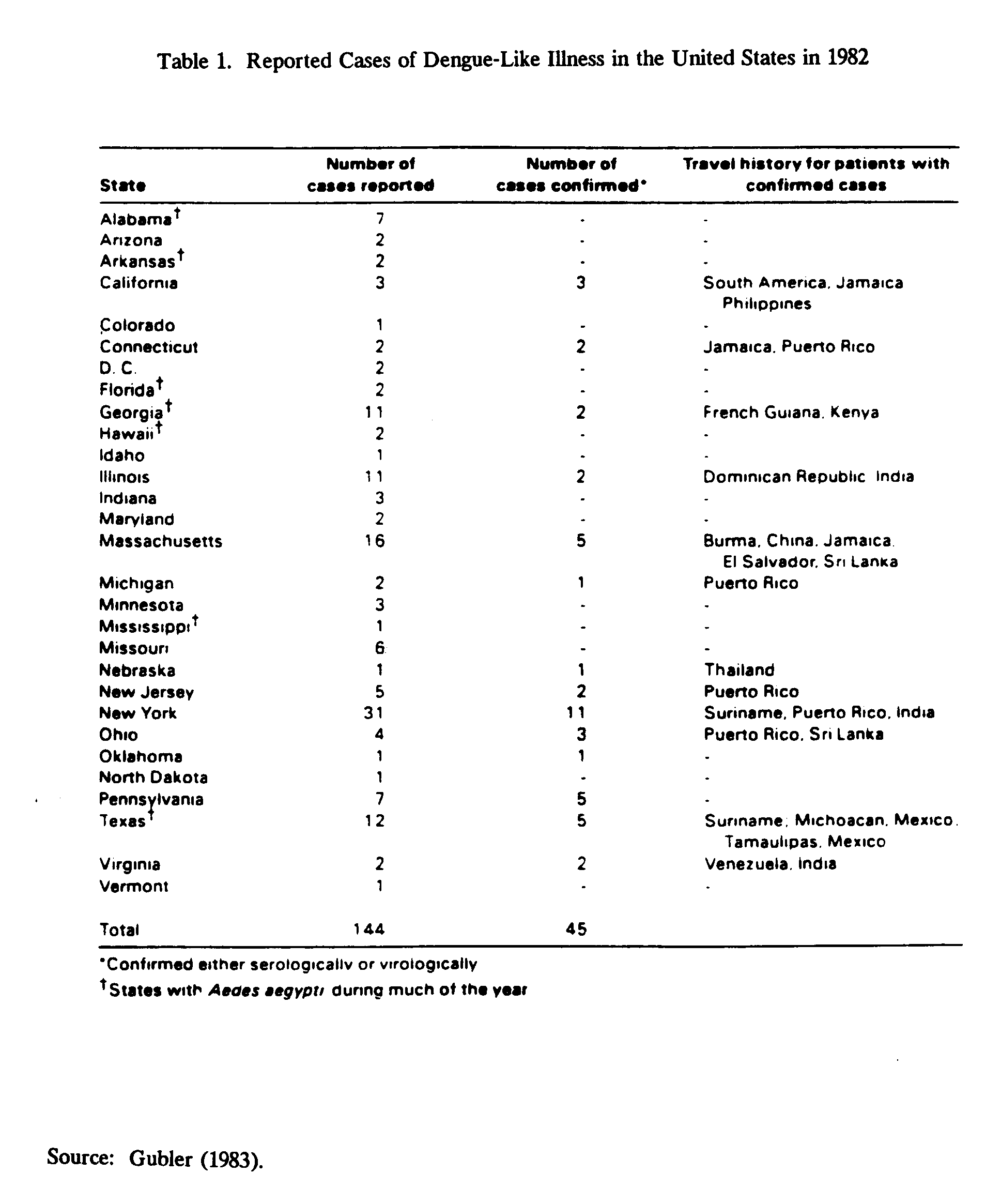{kind=link}
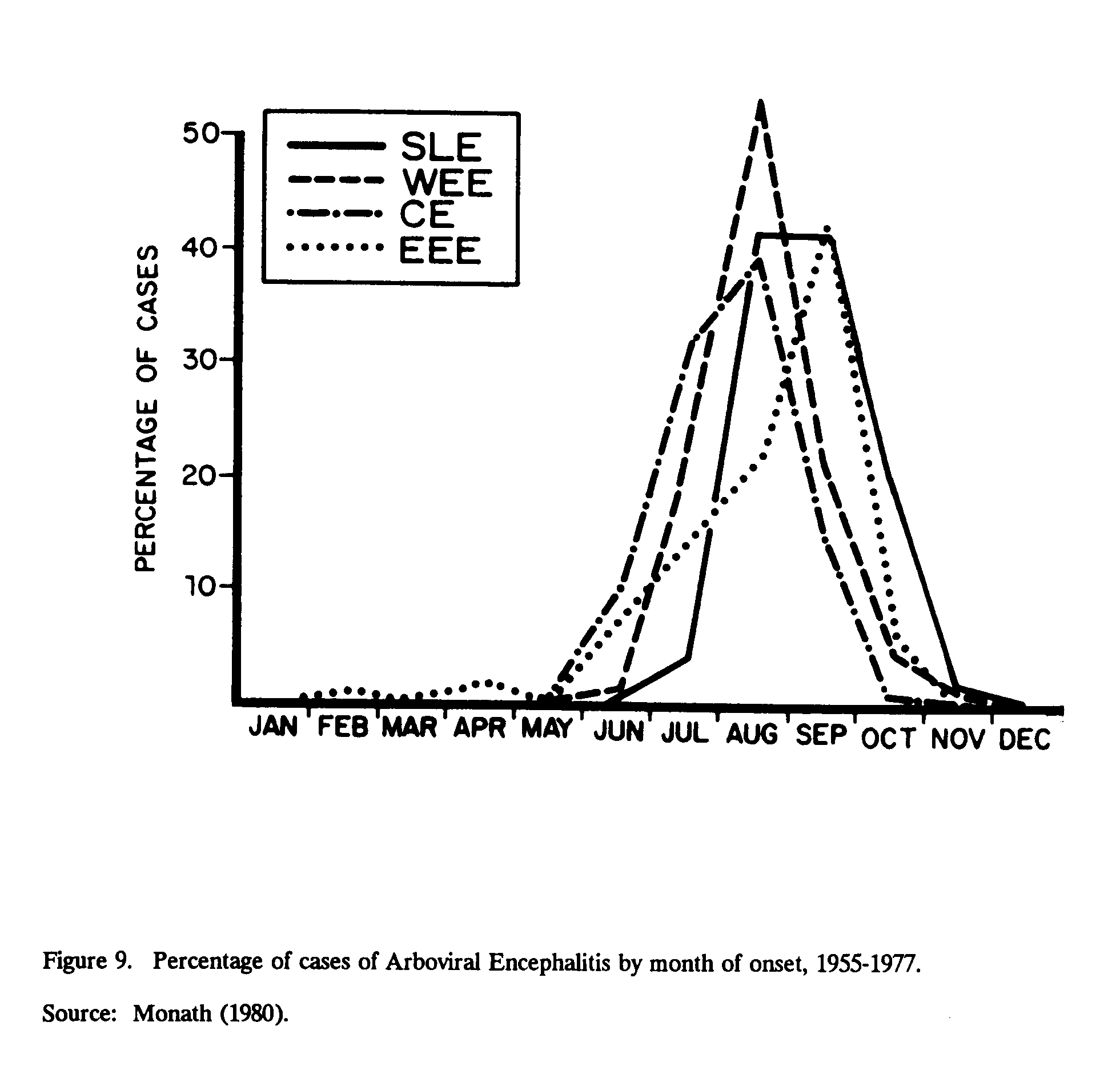{kind=link}
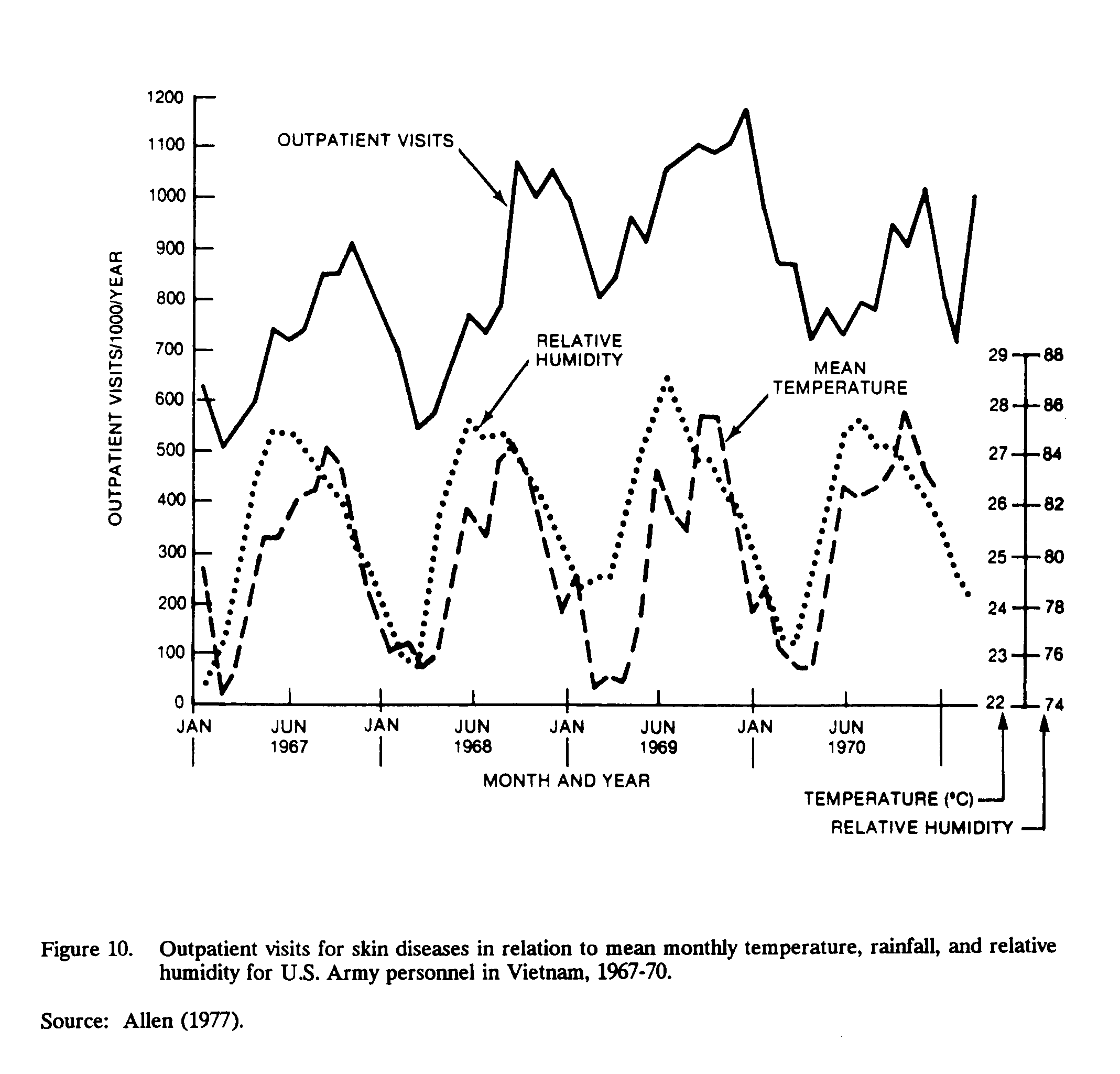{kind=link}