 Reproduced, with permission, from:
Diffey, B. L. 1991. Solar ultraviolet radiation effects on biological systems. Review in Physics in Medicine and Biology 36 (3): 299-328.
Reproduced, with permission, from:
Diffey, B. L. 1991. Solar ultraviolet radiation effects on biological systems. Review in Physics in Medicine and Biology 36 (3): 299-328.
Received 30 July 1990
1. Ultraviolet climatology
The Sun is responsible for the development and continued existence of life on Earth. We are warmed by the Sun's infrared rays and we can see with eyes which respond to the visible part of the Sun's terrestrial spectrum. More importantly, visible light is an essential component of photosynthesis, the process whereby plants, which are necessary for man's nutrition, derive their energy. However, the deleterious effects of sunlight on biological systems are due almost entirely to radiation within the ultraviolet spectrum of the Sun's emission.
The ultraviolet region of the electromagnetic spectrum is subdivided into three bands termed UVA, UVB and UVC. The subdivisions are arbitrary and differ somewhat depending on the discipline involved. Environmental photobiologists normally define the wavelength regions as: UVA, 400-320 nm; UVB, 320-290 nm; and UVC, 290-200 nm. The division between UVB and UVC is chosen as 290 nm since ultraviolet radiation (UVR) at shorter wavelengths is unlikely to be present in terrestrial sunlight, except at high altitudes (Henderson 1977). The choice of 320 nm as the division between UVB and UVA is perhaps more arbitrary. Although radiation at wavelengths shorter than 320 nm is generally more photobiologically active than longer wavelength UVR, recent advances in molecular photobiology indicate that a subdivision at 330-340 nm may be more appropriate (Peak and Peak 1986).
1.1. Atmospheric ozone
The quality and quantity of ultraviolet radiation at the Earth's surface depend on the energy output of the Sun and the transmission properties of the atmosphere. From a biological viewpoint, UVB radiation is by far the most significant part of the terrestrial ultraviolet spectrum and the levels of radiation in this waveband reaching the surface of the Earth are largely controlled by ozone, a gas which comprises approximately one molecule out of every two million in the atmosphere.
Ozone (O3) is created by the dissociation of oxygen (O2) by short wavelength UVR ( < 242 nm) in the stratosphere at altitudes between about 25 and 100 km. Absorption of UVR at wavelengths up to about 320 nm converts the O3 back to O2 and O (Chapman 1930). Dissociation of O3 is the mechanism responsible for preventing radiation at wavelengths less than about 290 nm from reaching the Earth's surface. In 1974 Molina and Rowland first warned that chlorofluorocarbons (CFCs) and other gases released by human activities could alter the natural balance of creative and destructive processes and lead to depletion of the stratospheric ozone layer. Substantial reductions of up to 50% in the ozone column observed in the austral spring over Antarctica and first reported in 1985 (Farman et al 1985) are continuing (SORG 1990). Coupled with this there has been a statistically significant downward trend in wintertime total ozone over the northern hemisphere of about 2-3% per decade for the past 30 years, although summertime ozone levels have remained approximately constant (Frederick 1990). In its report in June 1990, the UK Stratospheric Ozone Review Group concluded that there are serious limitations in our understanding and ability to quantify ozone depletion at the present levels of contaminant release and in our ability to predict the effects on stratospheric ozone of any further increases (SORG 1990).
1.2. Factors affecting terrestrial UVR
The spectral irradiance of UVR at the Earth's surface is modified by temporal, geographical and meteorological factors (Frederick et al 1989). In the ultraviolet, spectral irradiance falls by a factor of only two or three as the wavelength decreases from 400 to 320 nm at solar altitudes higher than 20deg., and then drops rapidly by three orders of magnitude or more from 320 to 290 nm as absorption by stratospheric ozone becomes important (figure 1).
1.2.1. Time of day. About 20-30% of total daily UVR is received one hour either side of midday in summer, with 75% between 9 am and 3 pm (table 1).
1.2.2. Season. In temperate regions the biologically damaging UVR reaching the Earth's surface shows strong seasonal dependence. However, seasonal variation is much less nearer the equator (table 2).
1.2.3. Geographical latitude. Annual UVR flux decreases with increasing distance from the equator. Very roughly, the annual number of minimal erythema doses (MED) on an unshaded, horizontal surface at mid-latitudes (20-60deg.) can be estimated as (Diffey 1990)
Annual MED = 2 x 10[4] exp(-latitude/20)
1.2.4. Clouds. Clouds reduce solar irradiance at the Earth's surface although changes in the ultraviolet region are not as great as those of total intensity, since water in clouds attenuates solar infrared much more than UVR. The risk of overexposure may be increased under these conditions because the warning sensation of heat is diminished.
Light clouds scattered over a blue sky make little difference to UVR intensity unless directly covering the sun, whilst complete light cloud cover reduces terrestrial UVR to about one half of that from a clear sky. Even with heavy cloud cover the scattered ultraviolet component of sunlight (often called skylight) is seldom less than 10% of that under clear sky (Paltridge and Barton 1978). However, very heavy storm clouds can virtually eliminate terrestrial UVR even in summertime (Diffey 1988).
1.2.5. Surface reflection. Reflection of UVR from ground surfaces, including the sea, is normally low (<7%). However gypsum sand reflects about 25% of incident UVB and fresh snow about 30% (Doda and Green 1980, 1981), although other authors (McCullough 1970, Blumthaler and Ambach 1985) have reported that the UVB reflectance of fresh snow exceeds 80%.
1.2.6. Altitude. In general, each 1 km increase in altitude increases the ultraviolet flux by about 6% (Cutchis 1980). Conversely, places on the Earth's surface below sea level are relatively poorer in UVB content than nearby sites at sea level. This is strinkingly apparent around the Dead Sea in Israel, 400 nm below sea level (Kushelevsky and Slifkin 1975).
1.3. Solar dosimetry in photobiology
The effects of UVR on biological systems show a strong dependence on wavelength. For this reason the specification of solar exposure in radiometric units is of limited value in many instances in photobiology because no account is taken of the spectral content of the source. Instead, a biologically effective exposure is often determined, which is obtained by weighting solar spectral irradiance by the appropriate function of wavelength (termed the 'action spectrum', see section 2.2) and integrating over the wavelength interval for which the action spectrum is non-zero. Biologically effective exposures can be determined in two ways (Diffey 1986): (i) by measuring the spectral irradiance of sunlight at one or more times during the period of exposure and calculating the weighted integral; or (ii) by using a device whose relative response to different wavelengths resembles the action spectrum for the particular photobiological effect.
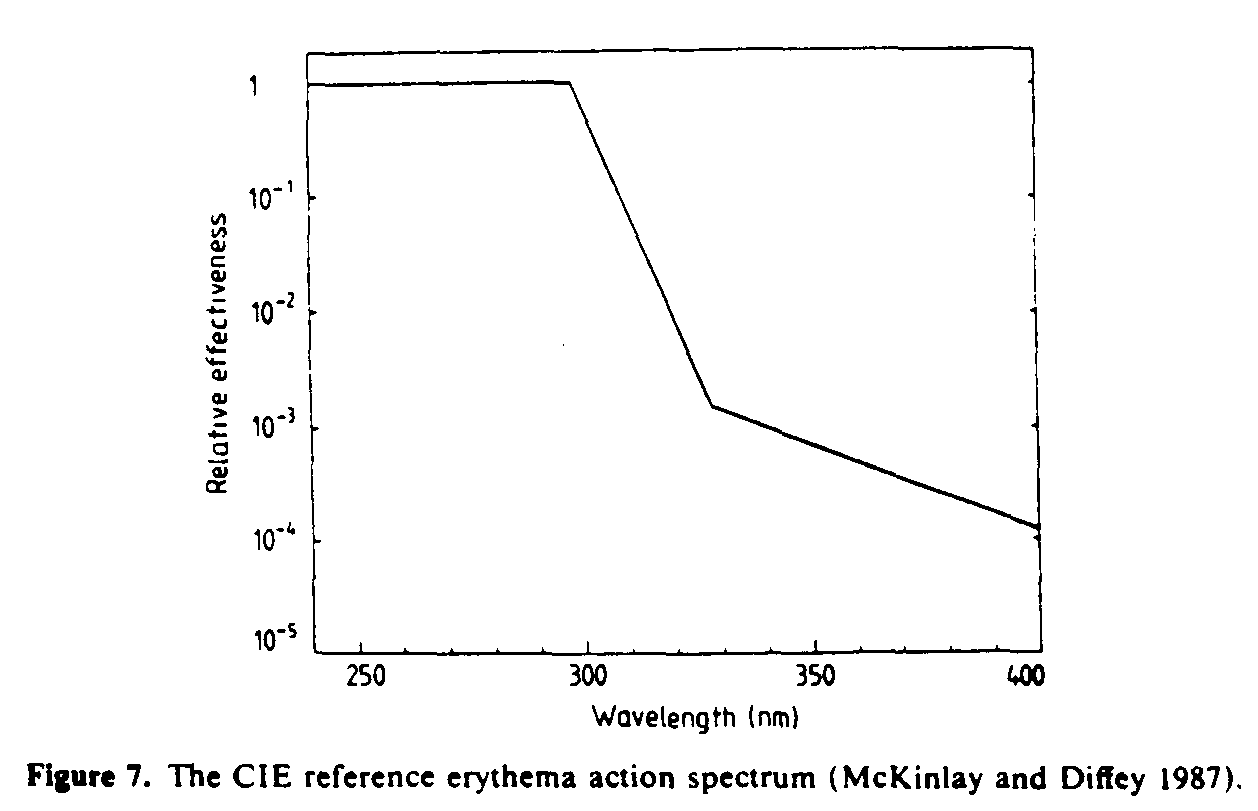
A criticism of much solar dosimetry in photobiology has focused on the mismatch that often occurs between spectral sensitivity of radiation detectors and the biological action spectra that they are designed to mimic (Calkins 1982). However, because only limited changes occur in the spectral power distribution of terrestrial sunlight, it has been shown that dosimetric errors that arise as a result of mismatched action spectra can be partly corrected for if detectors are calibrated using sunlight or simulated solar radiation as the source (Rupert 1982, Diffey 1987a). Radiation detectors used for monitoring solar UVR are commonly designed (Robertson 1972, Berger 1976) to have a spectral sensitivity which is a close match to the action spectrum for ultraviolet-induced erythema (see figure 7). The radiation dose recorded by these instruments is in units of 'minimal erythema dose', or MED, where one MED can be thought of as equivalent to that amount of solar exposure that produces a just perceptible reddening of unacclimatized white skin 24 h after exposure. Notwithstanding the difficulties of interpreting accurately the magnitude of such an imprecise unit, it has the advantage over radiometric units of indicating the biological consequences of the exposure.
1.4. Monitoring terrestrial UVR
In the 1950s Robertson in Australia developed an ultraviolet meter based upon the fluorescent conversion of UVB radiation to green light and which had a spectral sensitivity resembling the erythema action spectrum (Robertson 1972). The object of this work was to correlate the incidence of skin cancer in different parts of Australia with sunburning ultraviolet dose. The meter was subsequently redesigned by Berger in the USA (Berger 1976) as part of the Climatic Impact Assessment Program set up by the US Department of Transportation. Since 1973 a global network of these meters has been measuring half-hourly doses of sunburning UVR. From the original 10 stations, the network had increased by 1980 to almost 30 providing data on ultraviolet climatology at several sites around the World (Berger and Urbach 1982, Scotto et al 1988). This type of meter, generally referred to as the Robertson-Berger meter, has been used by other groups in the UK (Driscoll et al 1989), Switzerland (Blumthaler and Ambach 1990) and Kuwait (Kollias et al 1988). Data on monitoring solar UVB using other types of detector have been reported from Israel (Kushelevsky and Slifkin 1975), Holland (Schothorst et al 1987), Sweden (Wester 1987), Australia (Roy et al 1989) and New Zealand (Smith 1990). Monitoring of solar UVA has received little attention apart from measurements carried out in Scotland (Moseley et al 1983), although with the increasing importance of UVA effects on biological systems this may change in the future.
Spectral irradiance monitoring is not routinely carried out, presumably because of continual changes with solar elevation and the weather, and the high cost and complexity of equipment. An extensive set of spectral measurements was recorded at Davos in Switzerland by Bener in the 1960s, and these data, supplemented by theoretical calculations, provide a comprehensive tabulation of global spectral irradiance (Bener 1972). Spectroradiometric measurements of terrestrial UVR have also been published by workers in South Afrika (Kok 1972), the UK (Tarrant 1975, Diffey 1987a), Australia (Roy et al 1989) and Sweden (Wester 1984, Josefsson 1986).
1.5. Calculating terrestrial UVR
A number of computer models of terrestrial UVR have been developed (McCullough 1970, Green et al 1974, 1980, Green and Chai 1988, Diffey 1977, Björn and Murphy 1985, Bird and Riordan 1986, Paltridge and Barton 1978). Of particular note is the UVB Handbook compiled by Gerstl et al (1983) and which contains extensive tables and graphs of solar radiation from 290 to 320 nm in steps of 5 nm reaching the Earth's surface at any given location and specified time. Listings of computer programs for estimating terrestrial ultraviolet radiation are given in publications by Björn (1989) and Rundel (1986). Calculated results obtained from some of the published computer models are compared and discussed by Björn (1989).
2. Molecular and cellular ultraviolet photobiology
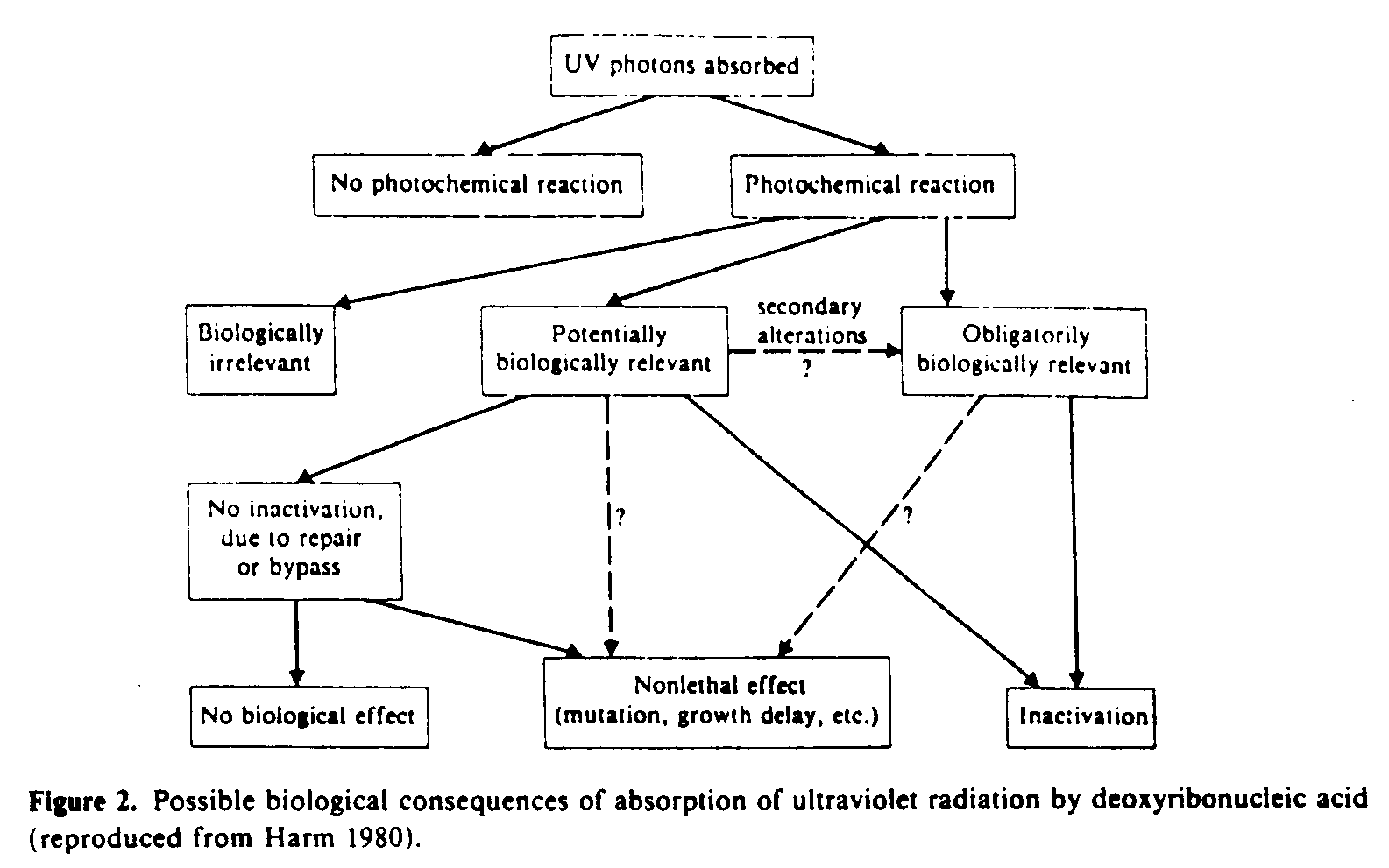
When biological species are irradiated with ultraviolet radiation a number of outcomes are possible (figure 2). Fortunately the vast majority of UV photons absorbed by nucleic acids (by far the most important molecules) do not cause lethality (inactivation).
2.1. Absorption characteristics of important biomolecules
The biological effects that result from ultraviolet irradiation are initiated by photochemical absorption by molecules of biological significance. The most important of these are the nucleic acids and to a much lesser extent the proteins and other molecules (Harm 1980).
2.1.1. Nucleic acids. It is the nucleotide bases that are the chromophores (absorbing centres) within nucleic acids. In deoxyribonucleic acid (DNA) these bases are the purine derivatives adenine and quanine, and the pyrimidine derivatives thymine and cytosine. Although the absorption spectra of the component bases differ slightly, their common features are an absorption maximum between 260-265 nm with a rapid reduction in absorption at longer wavelengths (figure 3).
2.1.2. Proteins. Like DNA, proteins absorb most strongly in the UVB and UVC regions with absorption maxima around 280 nm (figure 3). The absorbance of proteins is lower than that of nucleic acid solutions of equal concentration.
2.1.3. Other biomolecules. Absorption of UVR by other molecules can lead to biological consequences. These molecules include porphyrins, caratenoids, steroids and quinones.
2.2. Action spectra
An action spectrum is a plot of the relative effectiveness of radiation of different wavelengths to produce a given biological effect. The action spectrum for the formation of photoproducts might be expected to resemble the absorption spectrum of the molecules responsible for these photoproducts, although several conditions need to be fulfilled for this to be case (Jagger 1985). Nevertheless, comparison of experimentally determined action spectra with the absorption spectra of appropriate molesules can sometimes give insight into the molecule primarily responsible for the effect. For example, in 1928 Gates showed that the bactericidal action of UVR of different wavelengths in Staphylococcus aureus cells closely matched the absorption spectra of nucleotide bases. This observation was confirmed with other unicellular organisms (figure 4) and led to the realization that nucleic acids have a fundamental role in ultraviolet photobiology.
More recently, comparison of erythema action spectra, in patients with possible drug-induced photosensitivy, with the absorption spectra of suspect agents can confirm the diagnosis (Diffey and Farr 1988).
2.3. Photoproducts
The major photoproducts following the absorption of UVR by DNA are pyrimidine derivatives (Errera 1952). These photoproducts are cyclobutyl-type dimers (commonly called 'pyrimidine dimers' and by far the most significant), pyrimidine adducts, so-called 'spore photoproducts', pyrimidine hydrates and DNA-protein crosslinks (Patrick and Rahn 1976).
2.4. Inactivation of micro-organisms
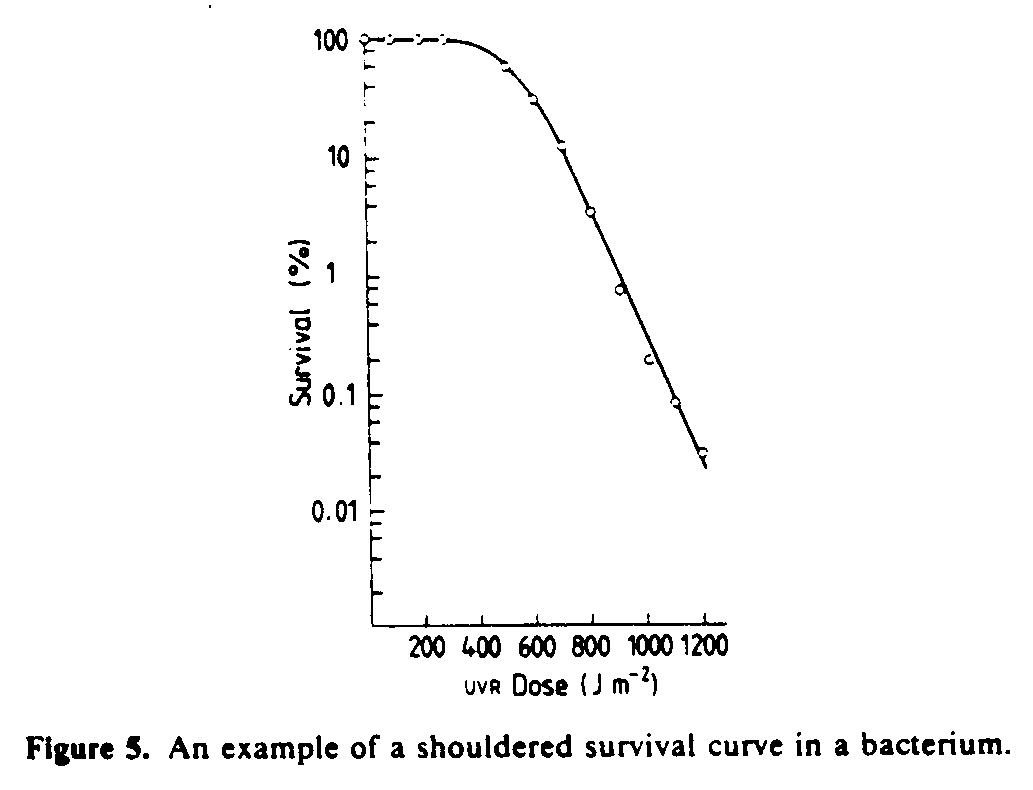
Cells and viruses can be inactivated by exposure to UVR; in other words, they lose their ability to reproduce. Inactivation is a stochastic process and is normally characterized by survival curves--a plot of the fraction of survivors as a function of UV dose. Mathematical treatment of survival curves has its basis in 'target theory', a concept familiar to radiobiologists (Wheldon 1988).
The simplest type of survival curve occurs when the number of survivors, S, decreases by a constant fractional amount for a given increment in exposure, i.e.
where So is the initial number of cells, D is the ultraviolet dose and is a factor that characterizes biological sensitivity. This simple model only holds if a single harmful event (hit) is sufficient to inactivate a cell. Often, however, inactivation of a single cell requires more than one hit and mathematical treatment of this condition leads to shouldered survival curves (Harm 1980) that are very often observed experimentally (figure 5).
2.5. Repair
Despite the fact that biological species, including man, are exposed to potentially harmful levels of solar UVR, mechanisms have evolved to protect cells and to repair damaged molecules.
The cell component most vulnerable to injury is nuclear DNA. A number of different DNA repair mechanisms have been established (Freifelder 1987), the best known being photoreactivation, excision repair, postreplication repair and sos repair.
2.5.1. Photoreactivation. This repair process begins with a photoreactivating enzyme binding to UV-induced pyrimidine dimers in the dark. If this complex is exposed to optical radiation between about 330 and 600 nm, the active enzyme separates from the dimer and a repaired DNA segment results.
2.5.2. Excision repair. This repair process takes place in the dark. Very simply, the defective region in one of the two helical strands of DNA is excised under enzymatic control and then subsequently replaced with normal nucleotides utilizing the complementary base-pairing information in the intact strand. Patients with the hereditary disorder, xeroderma pigmentosum, show reduced levels of excision repair which makes them prone to the development of multiple skin cancers from childhood onwards (Lehmann and Norris 1990).
2.5.3. Postreplication repair. UV-damaged DNA can replicate in such a way that gaps are left in the daughter strand opposite the damaged sites. Subsequently the gaps are filled by DNA synthesis, a process termed 'postreplication repair.'
2.5.4. sos repair. This mechanism of repair is not yet fully understood (Freifelder 1987) but is thought to include a bypass system that allows growth of the DNA chain across the damaged site of the thymine dimer. This is achieved at the cost of fidelity of replication and a great deal of evidence now indicates that sos repair is the major cause of ultraviolet-induced mutagenesis.
3. Effects of solar UVR on aquatic life
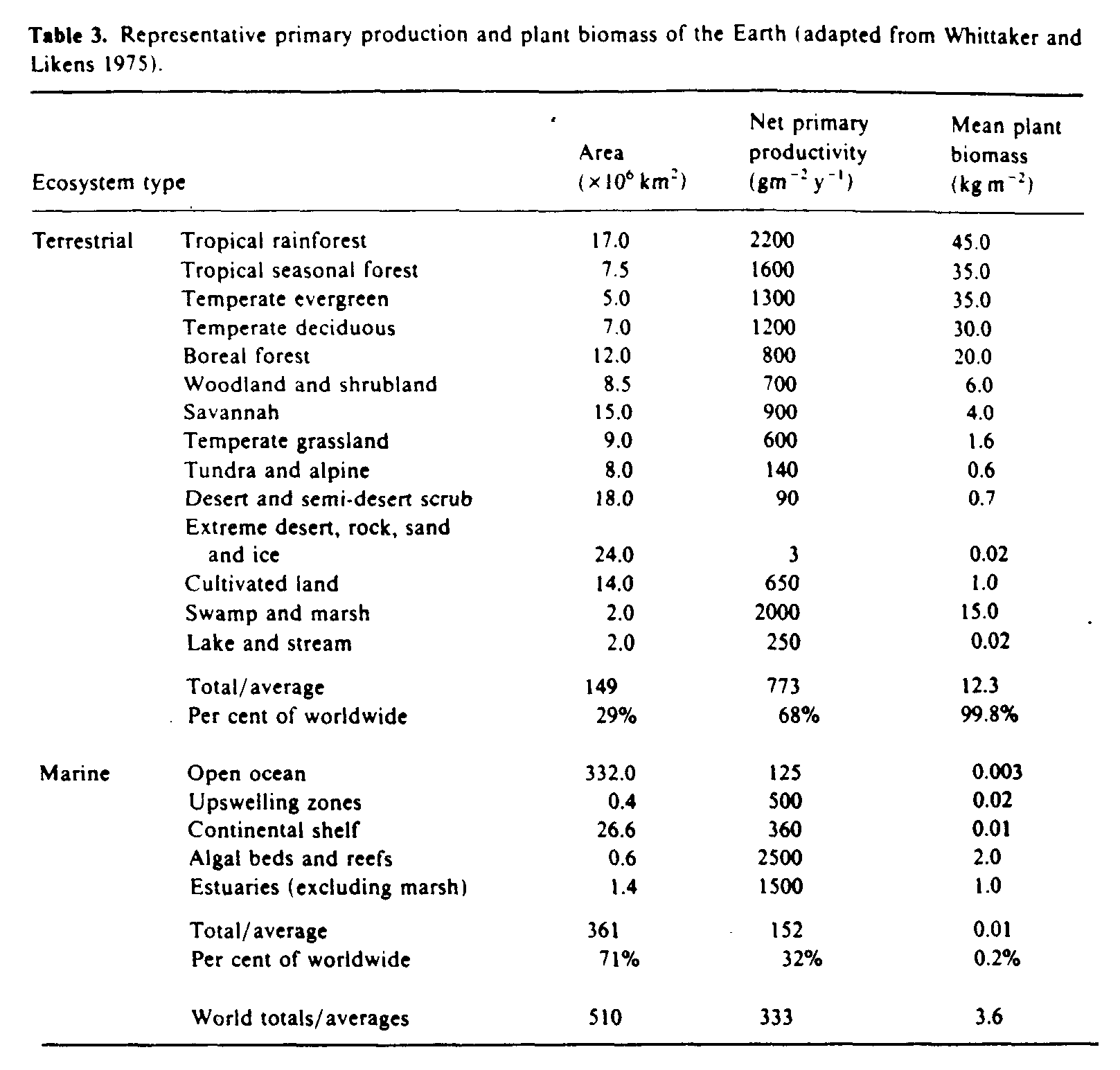
Marine waters cover 71% of the Earth's surface and contribute one third of global productivity (table 3). The organisms that live in either fresh water or the oceans can be classified into three groups; swimmers (nekton), bottom-dwellers (benthos) or drifters (plankton). Plankton are either phytoplankton (plants) or zooplankton (animals). Included in zooplankton are a group called icthyoplankton which are the drifting eggs and larvae of many species of fish.
Phytoplankton derive their energy from sunlight and as a consequence live in the upper 100 m of water where sufficient sunlight penetrates (termed the euphotic zone). Since zooplankton largely depend on phytoplankton as their primary source of food, they too reside in the euphotic zone.
The deleterious effects of solar UVR on aquatic organisms have been known for about 65 years (Huntsman 1925) and more recent studies have confirmed this (NAS 1979, 1984).
3.1. Penetration of solar UVR in natural waters
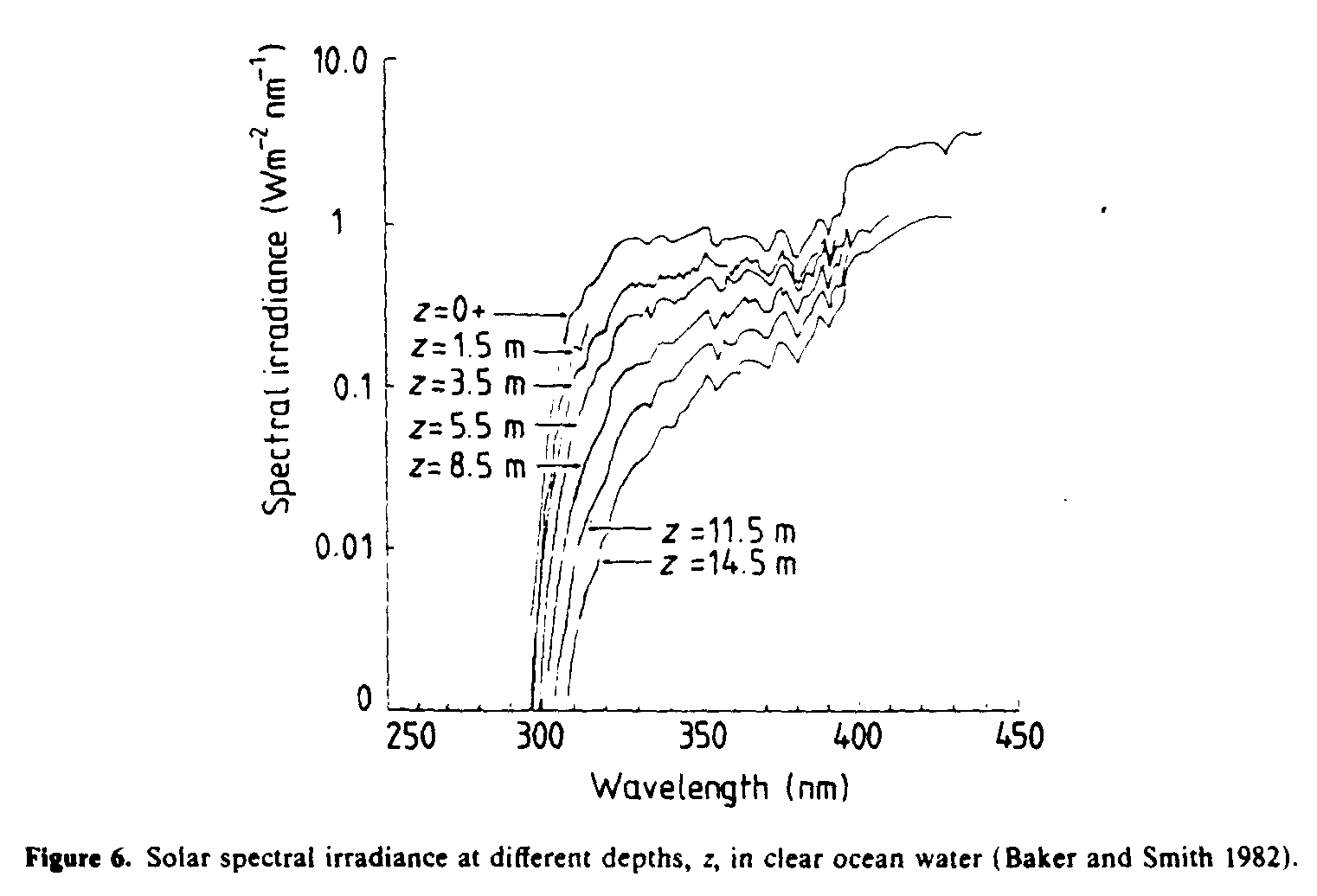
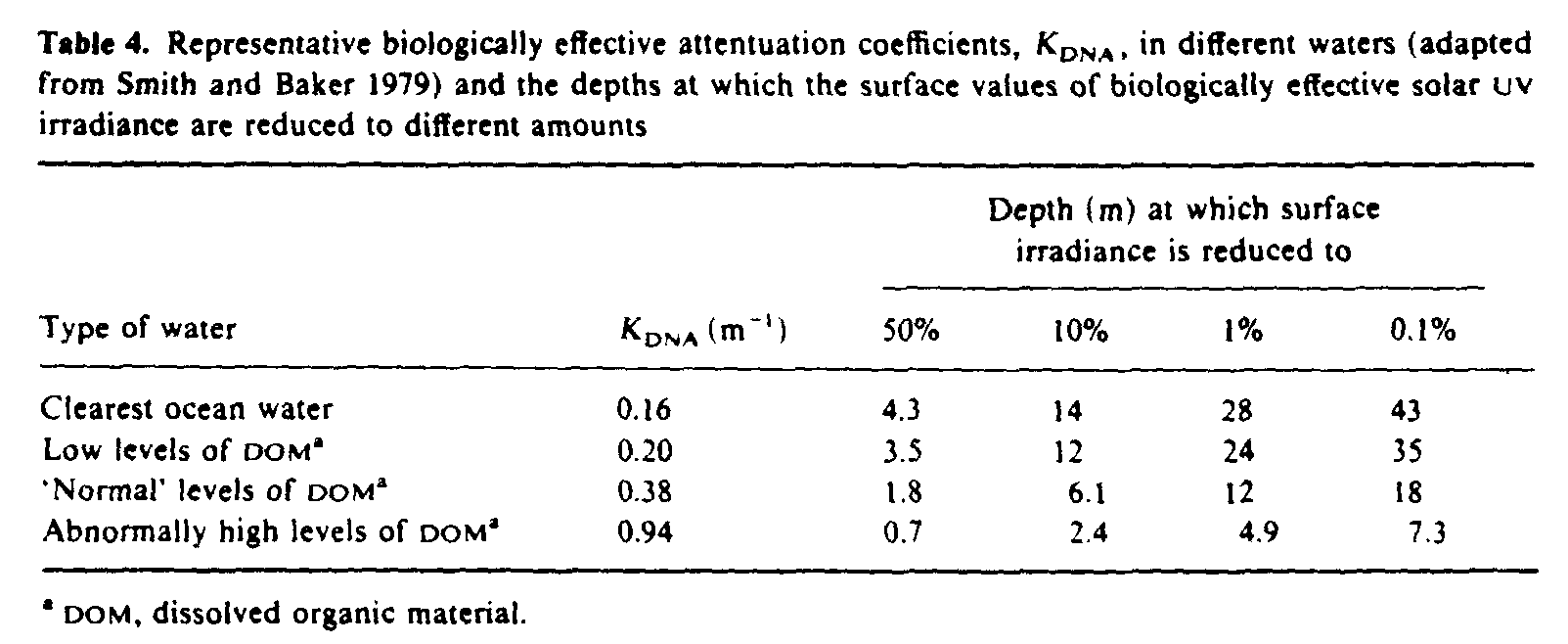
Solar UVR penetrates to biologically significant depths in natural waters, although the degree of penetration, particularly in the UVB region, depends strongly on the optical properties of the dissolved and suspended organic material. Spectral irradiances measured at different depths in clear ocean waters (Baker and Smith 1982) are shown in figure 6. By weighting spectral irradiance values at varying depths in different types of water with the action spectrum for causing damage in DNA (Setlow 1974) and integrating over the ultraviolet spectrum, Smith and Baker (1979) estimated biologically effective attenuation coefficients (table 4). The attenuation coefficients in this table are approximately numerically equal to the diffuse attenuation coefficients at 305 nm for all types of water (Smith and Baker 1979), and so the levels of penetration given in the last four columns of table 4 are a good guide to the penetration of UVB into sea water.
3.2. Effects on phytoplankton
Damaging effects of solar UVB on phytoplankton will occur at depths in excess of 20 m in clear waters and 5 m in cloudy waters (Worrest 1986). If it is assumed that phytoplankton sense and control their vertical position in such a way that limits UVB exposure to a tolerable level, then any increase in ambient UVB, as a result of ozone depletion, would necessitate a downward movement where there would be a corresponding reduction in light for photosynthesis, and hence a reduction in productivity (Smith 1989). Also, indirect effects of ambient levels of UVB radiation influence the survival of phytoplankton by decreasing their motility and inhibiting phototactic and photophobic responses (Worrest 1986). It has been estimated that a reduction of 25% in the ozone mantle would result in enhanced UVB levels at ocean surfaces that could lead to a 35% decrease in phytoplankton photosynthesis (Smith et al 1980), although it is important to recognize the uncertainty of this estimate.
3.3. Effects on zooplankton
Studies have shown that UVB irradiation of many zooplankton organisms will cause irrevesible damage and/or death, and decrease the fecundity of survivors (Damkaer 1982). Furthermore, it is believed that threshold UVB doses beyond those at which repair mechanisms are compromised, are near to the current ocean surface values for some organisms (Smith 1989). The threats to zooplankton from increased UVB are reduction in their food stocks (phytoplankton) and stresses at various states of egg and early larval developments, possibly causing mass mortalities leading to a decline in fish stocks (Chapman and Hardy 1988).
Fish comprise 18% of the average world animal protein intake, although in Asia, where more than half the world's population live, this figure is nearly 40% (Worrest 1986). Any increase in ambient UVB could result in a depletion of fish stocks as a food supply to humans, particularly if the species of phyto- and zooplankton that adapted to enhanced levels of UVB radiation were of different nutritional value from existing species.
4. Effects of solar UVR on plants
Twenty years ago virtually nothing was known about the effects of UVR on plants (Teramura 1986). Even today knowledge is principally limited to the effects on agricultural crops; little is known of the effects of UVR in other natural ecosystems such as forests, meadows, savannas, tundra and alpine areas (Tevini and Teramura 1989).
There are over 350 000 species of crop plants in the world and although 80 000 of these are edible, only about 3000 are harvested by man and used for food (Miller 1982). Of the 80 or so plant species that have been domesticated (Ehrlich et al 1977), just 15 species supply nearly all the food calories and three-quarters of the world's protein consumption (table 5).
The responses of plants to UV irradiation include physiological, biochemical, morphological and anatomical changes (table 6). In general UVR deleteriously affects plant growth, reducing leaf size and limiting the area available for energy capture. These findings have been achieved mainly through studies in greenhouses and exposure to artificial sources of ultraviolet radiation. Extrapolation to changes on crop yield as a result of increases in terrestrial solar UVR is difficult, and in those few field trials conducted outdoors the results were variable (Tevini and Teramura 1989). Furthermore, the effects of natural UVR on plants will be influenced by other stresses such as water shortage, mineral deficiency and increased concentrations of carbon dioxide. It is of note that increased ambient levels of CO2 (the greenhouse effect) have a beneficial effect on plants (Lemon 1983) but this may not necessarily compensate for the anticipated deleterious effects of increased ambient UVB as a consequence of ozone depletion.
Clearly more information is needed before a reliable assessment can be made on whether changes in ambient UVB are likely to affect significantly global crop productivity.
5. Effects of solar UVR on humans
The observable biological effects in man due to exposure from solar UVR are limited to the skin and to the eyes because of the low penetrating properties of UVR in human tissues. The penetration into skin is less than 1 mm (Bruls et al 1984) and UVR is absorbed by ocular tissues (mainly the cornea and the lens) before it reaches the retina.
5. 1. Effects of solar UVR on normal skin
The normal responses of the skin to UVR can be classed under two headings: acute effects and chronic effects. An acute effect is one of rapid onset and generally of short duration, as opposed to a chronic effect which is often of gradual onset and long duration. These effects should be distinguished from acute and chronic exposure conditions which refer to the length of the UVR exposure. The acute reactions considered will be sunburn, tanning and vitamin D production. Photo-ageing and skin cancer will be discussed as those chronic reactions produced by prolonged or repeated UVR exposure.
5.1.1. Sunburn. Sunburn, or erythema, is an acute injury following excessive exposure to solar UVR. The redness of the skin which results is due to an increased blood content of the skin by dilatation of the superficial blood vessels in the dermis, mainly the subpapillary venules. Some of the features of sunburn are discussed below.
5.1.1.1. Time course of sunburn. Half an hour of midday summer sunshine in the UK on the unacclimatized skin of Caucasian subjects is normally sufficient to result in a subsequent mild reddening of the skin. Following this degree of exposure erythema may not appear for about 4 h, although measurements using an instrument more sensitive than the eye at detecting erythema showed that vasodilatation begins to occur much sooner (Diffey and Oakley 1987). The erythema reaches a maximum at about 8-12 h after exposure and fades within 1 to 2 days (Olson et al 1966, Farr et al 1988). Exposing the skin for increasing periods to strong summer sunshine progressively shortens the time before the appearance of erythema, lengthens its persistence, and increases its intensity. High doses may result in oedema, pain, blistering and, after a few days, peeling.
5.1.1.2. Action spectrum for ultraviolet erythema. The effectiveness of UVR of different wavelengths in producing erythema has been determined repeatedly in a number of studies over the past 70 years (Diffey 1982). The technique is to determine the doses of UVR at a series of wavelengths necessary to produce a minimally perceptible redness 8 or 24 h after irradiation. This dose is termed the 'minimal erythema dose' or MED. The reciprocal of the MED is plotted against wavelength and the curve normalized to unity at the most effective wavelength. The MED at a given wavelength in a group of fair-skinned subjects are distributed lognormally. Studies in 254 normal subjects in the North East of England gave the median MED at 300 nm to be 34 mJ cm[-2] with a 95% confidence interval of 14-84 mJ cm[-2] (Diffey and Farr 1989).
Although the action spectra determined by various workers have shown differences, particularly in the spectral region 250-300 nm, there is good agreement that at wavelengths greater than 300 nm the effectiveness drops very rapidly, falling to an efficiency at 320 nm of about 1% of that at 300 nm. Recent studies (Parrish et al 1982, Gange et al 1986) have extended determination of the erythema action spectrum up to 400 nm and have shown that the erythemal effectiveness of UVR decreases with increasing wavelength through the ultraviolet spectrum, although the rate of change of effectiveness is much less from 330 to 400 nm, than from 300 to 330 nm.
A large number of published erythema action spectra were subject to statistical analysis and combined to produce the reference action spectrum (McKinlay and Diffey 1987) shown in figure 7. This action spectrum has been accepted by both the Commission Internationale de l'Eclairage (CIE) and the International Electrotechnical Committee (IEC), and has been shown to predict accurately the erythemal effectiveness of several polychromatic light sources differing greatly in spectral composition (Urbach 1987).
Although it can be seen from figure 7 that UVA radiation is much less erythmogenic than UVB radiation--broadly speaking by a factor of 1000--the much higher UVA irradiance present in sunlight means that in summertime UVA radiation contributes about 15-20% to the sunburn reaction.
5.1.1.3. Factors influencing the development of sunburn. Skin colour is an important factor in determining the ease with which the skin will sunburn. Whereas fair-skinned people require only about 15-30 min of midday summer sunshine to induce an erythremal reaction, people with moderately pigmented skin may require 1-2 h exposure and those with darkly pigmented skin (i.e. Negroes) will not normally sunburn. Other phenotype characteristics that may influence the susceptibility to sunburn are hair colour, eye colour and freckles (Azizi et al 1988, Andreassi et al 1987). Based on a personal history of response to 45-60 min of exposure to midday summer sun in early June (Fitzpatrick 1975), individuals can be grouped into six sun-reactive skin types (table 7).
There are anatomical differences in erythemal sensitivity. The face, neck and trunk are two to four times more sentitive than the limbs (Olson et al 1966). These anatomical differences are compounded by the variations in solar exposure on different parts of the body table 8). Vertical surfaces of an upright person receive about one half of the ambient UVR, whereas horizontal surfaces, such as the epaulet region of the shoulder, receive up to 75%.
There is no difference in sunburn susceptibility between sexes, although erythemal sensitivity may change with age in that young children and elderly people are said to be more sensitive (Hawk and Parish 1982). However, recent studies of erythemal sensitivity in children and elderly subjects have not confirmed this (Cox et al 1990).
Heat, humidity and wind have been shown to alter the erythemal sensitivity of mice exposed to artificial UVB radiation, but the significance of these atmospheric conditions upon the induction of sunburn in humans has not been clearly identified.
5.1.1.4. Sunburn and epidermal hyperplasia. In addition to erythema and tanning (section 5.1.2), thicknening (hyperplasia) of the epidermis is a significant component of a mild sunburn reaction. A single moderate exposure to UVB can result in up to a three-fold thickening of the stratum corneum within one to three weeks, and multiple exposures every one to two days for up to seven weeks will thicken the stratum corneum by about three- to five-fold (Miescher 1930). Skin thickness returns to normal about one to two months after ceasing irradiation.
Thickening of the skin, especially of the stratum corneum, after sun exposure can lead to a significant increase in protection against UVR by a factor of five or even higher. In Caucasians skin thickening is probably more important than tanning in providing endogenous photoprotection, although in darkly pigmented races it is likely that skin pigmentation is the most important means of protection against solar UVR.
5.1.2. Tanning. A socially desirable consequence of exposure to unfiltered sunlight is the delayed pigmentation of the skin known as tanning, or melanin pigmentation. Melanin pigmentation of skin is of two types: (i) constitutive--the colour of the skin seen in different races and determined by genetic factors only; and (ii) faculative--the reversible increase in tanning in response to solar UVR (and other external stimuli). The biology of the melanin pigmentary system is complex but for a good review see Fitzpatrick et al (1983).
5.1.2.1. Immediate pigment darkening (IPD). This is a transient darkening of exposed skin which can be induced by UVA and visible radiation (Rosen et al 1990). In general, the greater the constitutive tan, the greater is the ability to exhibit IPD. Immediate tanning can become evident within 5-10 min of exposure to summer sun and normally fades within 1-2 h.
Recent electron microscopic studies suggest that melanin photochemistry is the predominant mechanism of IPD (Beitner and Wennersten 1985, Honigsmann et al 1986). The function of IPD has not yet been demonstrated.
5.1.2.2. Delayed tanning. The more familiar delayed tanning becomes noticeable about one to two days after sun exposure, gradually increases for several days and may persist for weeks or months.
Following solar UVR exposure there is an increase in the number of functioning melanocytes and activity of the enzyme tyrosinase is enhanced (Fitzpatrick et al 1983). This leads to the formation of new melanin and hence an increase in the number of melanin granules throughout the epidermis.
Although a tanned skin does confer a degree of photoprotection, such protection seems to be no more than moderate, a factor of only two to three being achieved by a deep UVA-induced tan in the absence of skin thickening (Kaidbey and Kligman 1978). Melanin is not an effective sunscreen for Caucasian skin and it has been suggested (Morison 1985) that, contrary to popular belief, melanin is not an evolutionary adaptation to protect humans from the damaging effects of sunlight. Instead it is postulated that hominids developed melanin as a camouflage and as a device to keep their bodies warm in a forest environment.
5.1.2.3. Action spectrum for delayed tanning. The doses of UVR at different wavelengths necessary to induce delayed tanning were determined by Parrish et al ( 1982) for subjects with sun reactive skin types I and II (poor tanners) and by Gange et al (1986) for subjects who tan well (skin types III and IV). The action spectra obtained corresponded broadly with the erythema action spectrum (figure 7). The threshold doses at all wavelengths for erythema and pigmentation were similar for poor tanners, whereas in those subjects who are genetically capable of tanning easily the melanogenic doses in the UVA region were approximately one quarter of the doses required to produce a minimal erythema. That melanogenesis can be stimulated in individuals who tan well with suberythemal doses of solar UVR has been confirmed by field studies using unfiltered and optically filtered sunlight (Pathak and Fanselow 1983).
5.1.3. Production of vitamin D3. The only thoroughly established beneficial effect of solar ultraviolet radiation on the skin is the synthesis of vitamin D3. Solar radiation in the UVB waveband photochemically converts 7-dehydrocholesterol in the epidermis to previtamin D3. This previtamin immediately isomerizes to vitamin D3 in a reaction controlled by skin temperature and which takes two to three days to reach completion. Previtamin D3 is photolabile and excessive exposure to sunlight causes its photolysis to biologically inert photoproducts, lumisterol and tachysterol. In fact, production of previtamin D3 is limited to no more than 5-15% of the total 7-dehydrocholesterol content in the skin, no matter how long a person is exposed to sunlight. Once vitamin D3 is made in the skin it enters the blood for transport to the liver to be metabolized to 25-hydroxyvitamin D (Webb and Holick 1988). If vitamin D3 does not enter the circulation before sun exposure the following day, it can be rapidly degraded in the skin by sunlight to suprasterol 1, suprasterol 2 and 5,6-transvitamin D3--products which are believed to be biologically inert (Webb et al 1989). Thus sunlight, through its photochemical activity, is able to regulate the production of both previtamin D3 and vitamin D3 in the skin.
Only short exposures to sunlight are required to synthesize vitamin D3 in the skin; from spring until autumn 15 min exposure to the hands, arms and face between 9 am and 4 pm is adequate to provide our vitamin D3 requirement. However, in the UK it is likely there is insufficient ambient UVB during the winter to synthesize vitamin D3 (Webb et al 1988). Furthermore, increased melanin pigmentation in the skin can limit the production of vitamin D3 (Clemens et al 1982), as can increasing age (MacLaughlin and Holick 1985). It is not surprising, therefore, that the seasonal variation of solar UVB, and hence plasma 25-hydroxyvitamin D3 levels (Stamp and Round 1974), has led to calcium imbalance in the elederly (Exton-Smith et al 1972, Preece et al 1975) and nutritional osteomalacia in Asian immigrants to the UK (Dent et al 1973, Swan and Cooke 1971).
5.1.4. Photo-aging. The clinical signs of a photo-aged skin are dryness, deep wrinkles, accentuated skin furrows, sagging, loss of elasticity, mottled pigmentation and telangiectasia (Leyden 1990). These characteristics reflect profound structural changes in the dermis (Kligman and Kligman 1986). It has been speculated (Leyden 1990) that perhaps as much as 80% of solar UV-induced photo-aging occurs within the first 20 years of life, with the exception of those whose occupation or life style results in extensive exposure as adults.
5.1.4.1. Action specrum for photo-aging. The relative importance of different wavelengths in aging human skin cannot be readily determined because of the long latent period and slow evolution of photo-aging. Instead, extrapolation from experiments using hairless mice (Bissett et al 1989) or the miniature pig (Fourtanier and Berrebi 1989) is relied upon.
Since approximately one third of UVA radiation and less than 10% of UVB radiation incident on white skin penetrates to the dermis (Bruls et al 1984), it is not surprising that results from recent animal studies (Bissett et al 1989) have shown that chronic UVB and UVA irradiation in hairless mouse skin both result in histological, physical and visible changes characteristic of photo-aging. UVB radiation was only 20-50 times more efficient than UVA; this is in marked contrast to sunburn, suntan and non-melanoma skin cancer where UVB is about 1000 times more effective than UVA.
It should be remembered that solar radiation includes not only UVR but also visible and infrared radiation. Visible light is thought to be unimportant in photo-aging (Kligman and Kligman 1986) but studies have confirmed that infrared radiation can certainly damage the dermal matrix (Kligman and Kligman 1984).
5.1.4.2. Prevention of photo-aging. The application of topical sunscreens has been shown to inhibit photo-aging in mice chronically exposed to simulated solar radiation. Even products with a sun protection factor of two were shown to be effective (Young et al 1988), and it has been argued (Young 1990) that the incorporation of sunscreens into products used for daily cosmetic care could markedly delay the onset of photo-aging, as well as reduce the risk of skin cancer. Now that the importance of UVA in photo-aging is recognized, it will be important to use sunscreens that incorporate ingredients which provide photoprotection against both UVB and UVA (Honigsmann 1989).
5.1.4.3. Reversal of photo-aging. Photo-aged skin has the capacity to repair UVR-induced connective tissue damage (Kligman and Kligman 1986) and it has been shown that topical retinoic acid can enhance this process and improve photo-aged skin (Weiss et al 1988, Goldfarb et al 1990).
5.1.5. Non-melanoma skin cancer. Skin cancer is the most common human cancer and there is little dispute that chronic exposure to solar ultraviolet radiation is the most important cause of non-melanoma skin cancers (NMSC). The two common types of NMSC are basal cell carcinoma (BCC) and squamous cell carcinoma (SCC). BCC accounts for about 80% of all NMSC. The evidence for the carcinogenic effects of solar UVR is discussed below.
5.1.5.1. Anatomical distribution. Numerous studies have shown that about 90% of all BCC and more than half of all SCC occur on the head and neck (Urbach 1982). Studies made comparing the geometry of insolation of the head and neck areas with sites of NMSC show a close correlation (Urbach 1969, Diffey et al 1979).
5.1.5.2. Racial differences. Caucasians are much more likely to develop NMSC than races with more marked pigmentation. Furthermore, when skin cancer does occur in pigmented races it is not found predominantly on light exposed areas.
5.1.5.3. Phenotype. Genetic factors associated with a tendency to develop skin cancer are light eyes, fair complexion, light hair colour, tendency to sunburn and poor ability to tan (Urbach et al 1974).
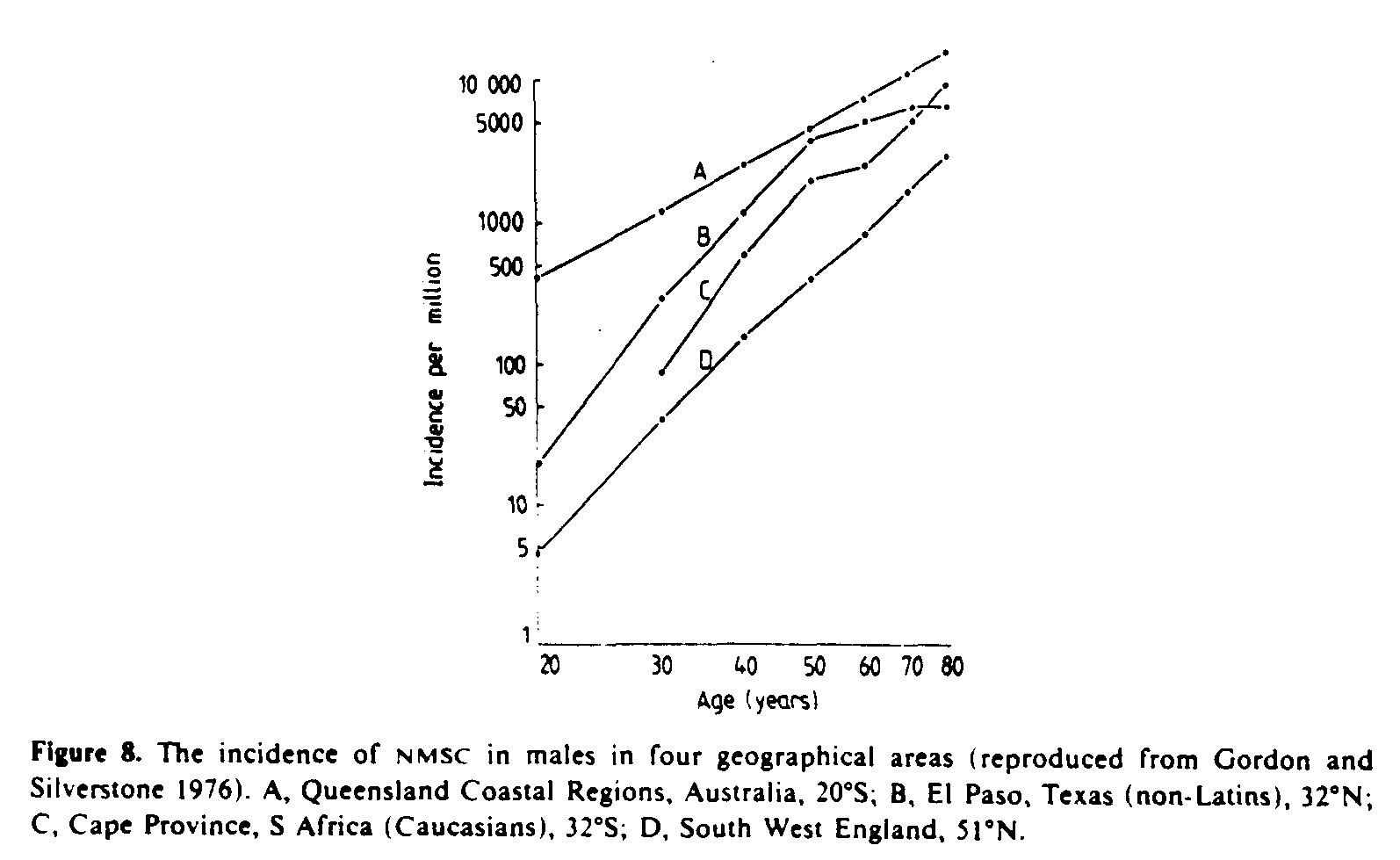
5.1.5.4. Geographical distribution. Surveys of the incidence of skin cancer carried out in various countries yield ample evidence that a geographical latitude gradient exists (Gordon and Silverstone 1976, Scotto et al 1981). Very roughly, the incidence doubles for every 10deg. decrease in latitude, provided that the population is genetically equally susceptible. The incidence of NMSC in four geographically different areas is illustrated in figure 8. Note that there is an approximate power law relationship between incidence and age with an exponent of approximately 4 to 5 indicating that NMSC is essentially a disease of the elderly.
5.1.5.5. Occupation. Epidemiological studies have shown that people who work outdoors are more likely to develop skin cancer than indoor workers (Swanbeck and Hillstrom 1971). Data collected in Sweden (Larkö and Swanbeck 1982) indicate that outdoor workers are about three times more likely to develop skin cancer than indoor workers.
5.1.5.6. The action spectrum for NMSC. Knowledge of the action spectrum for photocarcinogenesis is a prerequisite for assessing the risk of skin cancer from changes in the quality and quantity of solar UVR resulting from possible depletion of the ozone layer.
Early investigators (Wetzler 1959, Freeman 1975) exposed the ears of albino mice to narrow wavebands from an irradiation monochromator at wavelengths up to 320 nm and concluded that the action spectra for photocarcinogenesis and acute effects were similar. More recently, the hairless mouse has been used as the animal model (Cole et al 1986). In a series of careful investigations using this model and sources with seven different emission spectra, a group from Utrecht has published the most reliable action spectrum for photocarcinogenesis to date (Sterenborg and van der Leun 1987). This action spectrum is compared in figure 9 with the reference erythema action spectrum. It can be seen that for wavelengths longer than 290 nm (relevant for solar exposure) there is reasonable agreement between the two curves.
5.1.5.7. The risk of NMSC from sunlight. Application of multivariate analysis to the epidemiology of skin cancer has shown that, for a group of subjects with a given genetic susceptibility, age and environmental ultraviolet exposure are the two most important factors in determining the relative risk (Fears et al 1977). Other epidemiological studies have confirmed these findings, and this has led to a simple power law relationship which expresses the cumulative risk in terms of these factors (Schothorst et al 1985):
The symbols and
are numerical constants associated with the age dependence of the cumulative incidence and the biological amplification factor respectively.
Equation (3) is applicable to situations where the annual exposure received by an individual remains unaltered throughout life. In most instances changes in lifestyle with age mean that the annual UV exposure does not remain constant. For example, in adolescence and early adulthood sun-seeking holidays may be more popular than in middle and late age.
The situation of abrupt changes in annual exposure was examined in a series of experiments with mice (de Gruijl 1982), and led Slaper and van der Leun (1987) to modify equation (3) to estimate the risk of NMSC at age, T, as
Values of the exponents and
are normally derived from surveys of skin cancer incidence and UV climatology. Fears et al (1977) found the age exponent
, to be 4.7 and 4.1 for non-melanoma skin cancer (in males and females respectively). They found corresponding values of 3.0 and 2.5 for
. More recently, results from a survey by the National Cancer Institute in the US during the period June 1977 to May 1978 (Scotto et al 1981) estimated
to be 5.7 and 4.9 for squamous cell and basal cell carcinomas respectively, and
to be 2.9 and 1.7 for these two cancers. From data on skin cancer incidence in England and Wales (OPCS 1981) the exponent
for the age dependence of the cumulative incidence (males and females combined) is estimated to be 5.3. In calculations of risk estimates exemplary values for
and
of 5 and 2 respectively are often adopted (van der Leun 1984, Schothorst et al 1985, Diffey 1987b).
Since the action spectra for photocarcinogenesis and erythema are similar at wavelengths greater than 290 nm (figure 9), carcinogenic-effective exposure is often expressed in units of minimal erythema dose (MED). An example of relative risk estimates calculated using equation (4) is given in table 9. These data indicate that the risk of NMSC increases with increasing recreational sun exposure, but more importantly sun exposure during childhood leads to higher risks that the same exposure later in life. The importance of minimizing sun exposure to children has been demonstrated by Stern et al (1986) who estimated, using a mathematical model similar to the one described here, that regular use of a high protection factor sunscreen during the first 18 years of life could reduce the lifetime incidence of NMSC by more than 70%.
5.1.6. Malignant melanoma. Malignant melanoma is a tumour derived from the pigment cells (melanocytes) of the skin. Unlike NMSC, melanomas have a marked tendency to metastasize. In 1945 the 5 year survival in the USA for people with melanoma was only 41% although by 1983 this had risen to 83%, presumably because of earlier detection and treatment (Rigel et al 1987). Unfortunately the improvement in prognosis is offset by a rapid rise in incidence so that the mortality rate continues to rise. In the USA melanoma incidence has almost doubled in the past decade (Rigel et al 1987), and in Sweden there was a five-fold increase in incidence between 1958 and 1984 (Beitner et al 1990). Changing patterns of sun exposure are believed to be an important factor in the continuing rise in incidence. Some of the features of melanoma incidence are summarized below.
5.1.6.1. Geographical distribution. In general there is an inverse relationship between melanoma incidence and latitude of residence (Muir et al 1987), although there are some inconsistencies. For example, in Europe the incidence is higher in Scandinavia than in Mediterranean countries, and a similar lack of latitude gradient was seen in a study in Western Australia (Holman et al 1980).
5.1.6.2. Racial differences. Melanoma is much more common in Caucasians than in heavily pigmented races despite the fact that the latter tend to live in sunnier climates. An obvious explanation for this finding is that the greater degree of epidermal melanin found in the skin of darker races protects melanocytes in the basal layer of the epidermis from the harmful effects of solar UVR.
5.1.6.3. Phenotype. The single most important risk factor for developing melanoma is the total number of naevi (moles) larger than 2 mm diameter (MacKie et al 1989). Blue eyes, blond or red hair, light complexion and tendency to sunburn are other risk factors (Evans et al 1988).
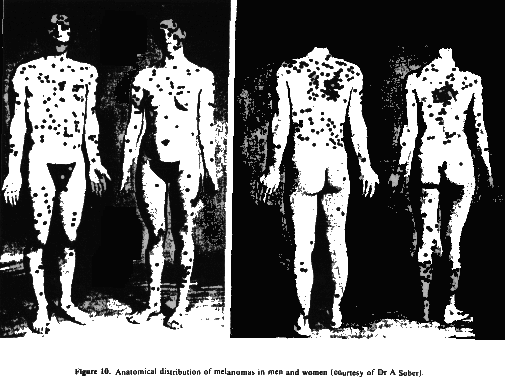
5.1.6.4. Anatomical distribution. Unlike NMSC which predominates on sites of highest insolation (head, neck and hands), melanoma occurs much more frequently on the trunk and legs (figure 10). This distribution lends support to the intermittent exposure hypothesis discussed below
5.1.6.5. Occupation and social status. Melanoma is more common in professional and technical indoor workers than in those who work outdoors, such as farmers (Armstrong 1988). Unlike all other cancers, melanoma is more likely in those of higher socioeconomic status (Vågerö et al 1990).
5.1.6.6. Migration and critical period studies. People born in Europe and who migrate to sunnier countries such as Israel, Australia or New Zealand after childhood have a risk of developing melanoma of about one quarter of that of people of European descent born in those countries (Lee 1989). However, arrival during childhood results in a comparable risk (Holman et al 1986). This observation suggests that sun exposure during childhood plays a crucial role in the aetiology of melanoma.
5.1.6.7. Intermittent exposure hypothesis. Although the evidence from epidemiological studies indicates an association between melanoma and sunlight exposure, it does not appear that cumulative sun exposure explains the relationship, as it does for NMSC. Instead, an intermittent exposure hypothesis proposes that infrequent intense exposure of unacclimatized skin to sunlight is related to increasing melanoma incidence and is more important than chronic sun exposure. This hypothesis is supported by the observation that most studies have shown that an increased risk of melanoma is associated with a past history of severe sunburn in childhood and adolescence (Armstrong 1988, MacKie and Aitchison 1982).
5.1.6.8. The action spectrum for melanoma. The action spectrum for the induction of melanoma by UVR is unknown. Although UVB is considered by some (Sober 1987, Koh et al 1990) to be the waveband primarily responsible, the possibility that the action spectrum lies within the UVA, visible or even infrared parts of the solar spectrum cannot, as yet, be discounted (Loggie and Eddy 1988).
5.2. Effects of solar UVR on the eye
The evidence linking solar UVR exposure to ocular damage has, until recently, been weak. However, epidemiological studies published within the past three years have strengthened this evidence and have shown that UVB is the most damaging part of the ultraviolet spectrum to the eye.
5.2.1. Effects on the cornea. The association between UVB exposure and photokeratitis, or snowblindness, has been established for some while ( Pitts and Tredici 1971). Ambient levels of solar UVR are such that this acute phototoxic corneal disorder is largely confined to snowfields and deserts. It has been estimated (Sliney 1987) that approximately 2 h exposure outdoors around noon on snow-covered terrain is sufficient to induce photokeratitis, and that an exposure of 6-8 h may be needed in sandy terrain.
It has been shown recently (Taylor et al 1989) that outdoor workers with high solar exposure have an approximate three-fold risk of developing pterygium (a fleshy growth on a normally clear cornea) and a six-fold risk of having climatic droplet keratopathy (a deposition of altered proteins on the superficial cornea leading to opacification).
5.2.2. Effects on the lens. Studies in experimental animals have confirmed that the development of certain types of cataract (opacity of the lens) are associated with ocular UVB exposure (Pitts et al 1976, Zigman et al 1974). A recent epidemiological study (Taylor et al 1988) has found a similar response in humans. The study examined 838 waterman who worked on the Chesapeake Bay, Maryland, USA (latitude 37deg.N), and showed a clear, consistent relationship between the development of cortical cataract and estimated ocular solar UVB exposure. A doubling of cumulative exposure increased the risk of cortical cataract by a factor of 1.6. No evidence was found for a threshold level of sun exposure either in terms of irradiance, duration of exposure or age. Also no association was found between nuclear cataracts and UVB exposure, nor between either cortical or nuclear opacities and UVA exposure.
5.2.3. Effects on the retina. A recent epidemiological study (West et al 1979) examined the incidence of senile macular degeneration in outdoor workers and found no association with solar UVB exposure.
A case control study (Tucker et al 1985) of intraocular malignant melanoma suggested that exposure to sunlight, especially in early life, may be important in the development of this condition.
It has been recognized that the macula in the aphakic eye may be at risk from solar UVR exposure following cataract surgery (Fishman 1986, Rosen 1986). The natural crystalline lens absorbs all incident UVB and almost all UVA radiation and concern has been expressed that some types of implanted thin plastic intraocular lenses may not provide adequate UV protection to the retina. As yet, no human data exist to confirm whether or not the retina is at risk (Taylor 1989).
References
Andraessi K, Simoni S, Fiorini P and Fiamiani M 1987 Phenotypic characters related to skin type and minimal erythema dose Photodermatology 4 43-6
Armstrong B K 1988 Epidemiology of malignant melanoma: intermittent or total accumulated exposure to the sun? J. Dermatol, Surg. Oncol. 14 835-49
Azizi E, Lusky A, Kushelevsky A P and Schewach-Millet M 1988 Skin type, hair colour, and freckles are predictors of decreased minimal erythema ultraviolet radiation dose J. Am. Acad. Dermatol. 19 32-8
Baker K S and Smith R C 1982 Spectral irradiance penetration in natural waters The Role of Solar Ultraviolet Radiation in Marine Ecosystems ed J Calkins (New York: Plenum) pp 233-46
Beitner H, Norell S E, Ringborg U, Wernersten G and Mattson B 1990 Malignant melanoma: aetiological importance of individual pigmentation and sun exposure Br. J. Dermatol. 122 43-51
Beitner H and Wennersten G 1985 A qualitative and quantitive transmission electron microscopie study of immediate pigment darkening reaction Photodermatology 2 273-8
Bener P 1972 Approximate values of intensity of natural ultraviolet radiation for different amounts of atmospheric ozone Final Technical Report DAJA 37-68-C-1017 (London: European Research Office, United States Army)
Berger D S 1976 The sunburning ultraviolet meter: design and performance Photochem. Photobiol 24 587-93
Berger D S and Urbach F 1982 A climatology of sunburning ultraviolet radiation Photochem. Photobiol. 35 187-92
Bird, R E and Riordan C 1986 Simple solar spectral model for direct and diffuse irradiance on horizontal and tilted planes at the earth's surface for cloudless atmospheres J. Climate Appl. Meteorol. 25 87-97
Bissett D L, Hannon D P and Orr T V 1989 Wavelength dependence of histological, physical, and visible changes in chronically UV-irradiated hairless mouse skin Photochhem. Photobiol. 50 763-9
Björn L O 1989 Computer programs for estimating ultraviolet radiation in daylight Radiation Measurement in Photobiology ed B L Diffey (London: Academic) pp 161-89
Björn L O and Murphy T M 1985 Computer calculation of solar ultraviolet radiation at ground level Physiol. Veg. 23 555-61
Blumthaler M and Ambach W 1985 Neure Messugen der Albedo verschiedener Oberflaechen fur erythemwirksame Strahlung Annal. Meteorol. 22 114-5
-----1990 Indication of increasing solar ultraviolet-BC radiation flux in alpine regions Science 248 206-8
Bruls W A G, Slaper H, van der Leun J C and Berrins L 1984 Transmission of human epidermis and stratum corneum as a function of thickness in the ultraviolet and visible wavelengths Photochem. Photobiol. 40 485-94
Calkins J 1982 Measuring devices and dosage units The Role of Solar Ultraviolet Radiation in Marine Ecosystems ed J Calkins (New York: Plenum) pp 169-79
Chapman S 1930 A theory of upper atmospheric ozone Mem. Roy. Meteorol. Soc. 3 103-25
Chapman J and Hardy J T 1988 Effects of middle ultraviolet radiation on marine fishes Final Report Oregon State University US EPA Coop. Argrmt. CR-812688-02-0
Clemens T L, Adams J S, Henderson S L and Holick M F 1982 Increased skin pigment reduces the capacity of skin to synthesize vitamin D, Lancet 1 74-6
Cole C A, Forbes P D and Davies R E 1986 An action spectrum for photocarcinogenesis Photochem. Photobiol. 43 275-84
Cox N H, Farr P M and Diffey B L 1990 The relationship between chronological age and erythemal response to UVB irradiation Br. J. Dermatol. 122 272-3
Cutchis P 1980 A formula for comparing annual damaging ultraviolet (DUV) radiation doses at tropical and mid-latitude sites Final Report FAA-EE 80-21 (Washington, DC: US Department of Transportation, Federal Aviation Administration Office of Environment and Energy)
Damkaer D M 1982 Possible influence of solar UV radiation in the evolution of marine zooplankton The Role of Solar Ultraviolet Radiation in Marine Ecosystems ed J Calkins (New York: Plenum) pp 701-6
Dent C E, Round J M, Rowe D J F and Stamp T C B 1973 Effect of chapattis and ultraviolet irradiation on nutritional rickets in an Indian immigrant Lancet i 1282
Diffey B L 1977 The calculation of the spectral distribution of natural ultraviolet radiation under clear day conditions Phys. Med. Biol. 22 309-16
-----1982 The consistency of studies of ultraviolet erythema in normal human skin Phys. Med. Biol. 27 715-20
-----1986 Possible errors involved in the dosimetry of solar UV-B radiation Stratospheric Ozone Reduction, Solar Ultraviolet Radiation and Plant Life R C Worrest and M M Caldwell (Berlin: Springer-Verlag) pp 75-86
-----1987a A comparison of dosimeters used for solar ultraviolet radiometry Photochem. Photobiol. 46 55-60
-----1987b Analysis of the risk of skin cancer from sunlight and solaria in subjects living in Northern Europe Photodermatology 4 118-26
-----1990 Human exposure to ultraviolet radiation Semin. Dermatol. 9 2-10
-----1988 unpublished data
Diffey B L and Farr P M 1988 The action spectrum in drug induced photosensitivity Photochem. Photobiol. 47 49-54
-----1989 The normal range in diagnostic phototesting Br. J Dermatol. 120 517-24
Diffey B L, Kerwin M and Davis A 1977 The anatomical distribution of sunlight Br. J. Dermatol. 97 407-10
Diffey B L and Oakley A M 1987 The onset of ultraviolet erythema Br. J. Dermatol. 116 183-7
Diffey B L, Tate T I and Davis A 1979 Solar dosimetry of the face: the relationship of natural ultraviolet radiation exposure to basal cell carcinoma localisation Phys. Med. Biol. 24 931-9
Doda D D and Green A E S 1980 Surface reflection measurements in the UV from an airborne platform. Part 1 Appl. Opt. 19 2140-5
-----1981 Surface reflection measurements in the UV from an airborne platform. Part 2 Appl. Opt. 20 636-42
Driscoll C M H, Whillock M J, Gall A, Clark I E, Pearson A J, Blackwell R P, Strong J C and McKinlay A F 1989 Solar radiation measurements at three sites in the UK, May 1988-April 1989 Report NRPB-M184 (Chilton: National Radiological Protection Board)
Ehrlich P R., Ehrlich A H and Holdren J P 1977 Eco-Science: Population, Resources, Environment (San Francisco, CA: Freeman) pp 283-385
Errera M 1952 Etude photochemique de l'acide désoxyribonucleique I. Mesures énergétiques Biochim. Biophys. Acta 8 30-7
Evans, R D, Kopf A W, Lew R A, Rigel D S, Bart R S, Friedman R J and Rivers J K 1988 Risk factors for the development of malignant melanoma-I: review of case-control studies J. Dermatol. Surg. Oncol. 14 393-408
Exton-Smith A N, Stanton B R and Windsor A C M 1972 Nutrition of housebound old people (London: King Edward's Hospital Fund for London)
Farman J C, Gardiner B G and Shaklin J D 1985 Large losses of total ozone in Antarctica reveal seasonal ClOx/NOx interaction Nature 315 207-10
Farr P M, Besag J E and Diffey B L 1988 The time-course of UVB and UVC erythema J. Invest. Dermatol. 91 454-7
Fears T R, Scotto J and Schneiderman M A 1977 Mathematical models of age and ultraviolet effects on the incidence of skin cancer among whites in the United States Am. J. Epidermiol. 105 420-7
Fishman G A 1986 Ocular phototoxicity: guidelines for selecting sunglasses Surv. Ophthalmol. 31 119-24
Fitzpatrick T B 1975 Soleil et peau J. Med. Esthet. 2 33-4
Fitzpatrick T B, Szabo G and Wick M W 1983 Biochemistry and physiology of melanin pigmentation Biochemistry and Physiology of the Skin ed L A Goldsmith (Oxford: Oxford University Press) pp 687-7l2
Food and Agriculture Organisation 1981 FAO Production Yearbook vol 35 (Rome: FAO)
Fourtainier A and Berrebi C 1989 Miniature pig as an animal model to study photoageing Photochem. Photobiol. 50 771-84
Frederick J E 1990 Trends in atmospheric ozone and ultraviolet radiation: mechanisms and observations for the Northern Hemisphere Photochem. Photobiol. 51 757-63
Frederick J E, Snell H E and Haywood E K 1989 Solar ultraviolet radiation at the earth's surface Photochem. Photobiol. 50 443-50
Freeman R G 1975 Data on the action spectrum for ultraviolet carcinogenesis J. Nat. Cancer Inst. 55 1119-21
Freifelder D 1987 Molecular Biology 2nd edn (Boston: Jones and Bartlett) pp 277-92
Gange R W, Park Y K, Auletta M, Kagetsu N, Blackett A D and Parrish J A 1986 Action Spectra for cutaneous responses to ultraviolet radiation The Biological Effects of UVA Radiation ed F Urbach and R W Gange (New York: Praeger) pp 57-65
Gates F L 1928 On nuclear derivatives and the lethal action of ultraviolet light Science 68 478-80
Gerstl S A W, Zardecki A and Wiser H L 1983 UVB Handbook vol 1 Los Alamos National Laboratory Document LA-IR-83-728
Goldfarb M T, Ellis C N and Voorhees J J 1990 Topical tretinoin: its use in daily practice to reverse photoaging Br J. Dermatol. 122 (Suppl. 35) 87-91
Gordon D and Silverstone H 1976 Worldwide epidemiology of pre-malignant and malignant cutaneous lesions Cancer of the Skin ed R Andrade (Philadelphia, PA: Saunders) pp 405-55
Green A E S and Chai S 1988 Solar spectral irradiance in the visible and infrared regions Photochem. Photobiol. 48 477-86
Green A E S, Cross K R and Smith L A 1980 Improved analytic characterization of ultraviolet skylight Photochem. Photobiol. 31 59-65
Green A E S, Swada T and Shettle E P 1974 The middle ultraviolet reaching the gound Photochem. Photobiol. 19 251-9
de Gruijl F R 1982 The dose-response relationship for UV tumorigenesis PhD Thesis University of Utrecht
Harm W 1980 Biological Effects of Ultraviolet Radiation (Cambridge: Cambridge University Press)
Hawk J L M and Parrish J A 1982 Responses of normal skin to ultraviolet radiation The Science of Photomedicine ed J D Regan and J A Parrrsh (New York: Plenum) pp 219-60
Henderson S T 1977 Daylight and its Spectrum (Bristol: Adam Hilger) p33
Holman C D J, Armstrong B K and Heenan P J 1986 The cause of malignant melanoma: results from the West Australian Lions melanoma research project Recent Results Cancer Res. 102 18-37
Holman C D J, Gibson I M, Stephenson M and Armstrong B K 1983 Ultraviolet irradiation of human body sites in relation to occupation and outdoor activity: field studies using personal UVR dosimeters Clin. Exp. Dermatol. 8 269-77
Holman C D J, Mulroney C D and Armstrong B K 1980 Epidemiology of preinvasive malignant melanoma in Western Australia Br. J. Cancer 25 317-23
Honigsmann H 1989 UVA and human skin J. Photochem. Photobiol. B Biol. 4 229
Honigsmann H G, Schuler W, Aberer N, Romani N and Wolff K 1986 Immediate pigment darkening phenomenon: A reevaluation of its mechanisms J. Invest. Dermatol. 87 648-52
Huntsman A G 1925 Limiting factors for marine animals I. The lethal effect of sunlight Contrib. Can. Biol. 2 83-8
Jagger J 1985 Solar-UV Actions on Living Cells (New York: Praeger) pp 174-6
Josefsson W 1986 Solar ultraviolet radiation in Sweden SMHI Reports: Meteorology and Climatology vol 53 (Norrkoping: Sweden Meteorological and Hydrological Institute) p 71
Kaidbey K H and Kligman A M 1978 Sunburn protection by longwave ultraviolet radiation-induced pigmentation Arch. Dermatol. 114 46-8
Kligman L H and Kligman A M 1984 Reflections on heat Br. J. Dermatol. 110 369-75
-----1986 The nature of photoageing: its prevention and repair Photodermatol. 3 215-27
Koh H K, Kligler B E and Lew R A 1990 Sunlight and cutaneous malignant melanoma: evidence for and against causation Photochem. Photobiol. 51 765-79
Kok C J 1972 Spectral irradiance of daylight for air mass 2 J. Phys. D: Appl. Phys. 5 185-8
Kollias N, Baqer A H and Sadiq 1 1988 Measurement of solar middle ultraviolet radiation in a desert environment Photochem. Photobiol. 47 565-9
Kushelevsky A P and Slifkin M A 1975 Ultraviolet measurements at the Dead Sea and Beersheba Isr. J. Med. Sci. 11 488-90
Larkö O, and Swanbeck G 1982 Is UVB treatment of psoriasis safe? A study of extensively UVB-treated psoriasis patients compared with a matched control group Acta Dermatol. Venereol, Stockholm 62 507- 12
Lee J A H 1989 The relationship between malignant melanoma of skin and exposure to sunlight Photochem. Photobiol. 50 493-6
Lehmann A R and Norris P G 1990 DNA repair deficient photodermatoses Semin. Dermatol. 9 55-62
Lemon E R 1983 CO2 and plants The Response of Plants to Rising Levels of Atmospheric Carbon Dioxide (Boulder, CO: Westview Press)
Leyden J J 1990 Clinical features of ageing skin Br J. Dermatol. 122 (Suppl. 35) 1-3
Loggie B W and Eddy J A 1988 Solar considerations in the development of cutaneous melanoma Semin. Oncol. 15 494-9
McCullough E C 1970 Qualitative and quantitative features of the clear day terrestrial solar ultraviolet radiation environment Phys. Med. Biol. 15 723-34
MacKie R M, Freudenberger T and Aitchison T C 1989 Personal risk-factor chart for cutaneous melanoma Lancet ii 487-90
McKinlay A F and Diffey B L 1987 A reference action spectrum for ultraviolet induced erythema in human skin Human Exposure to Ultraviolet Radiation: Risks and Regulations ed W F Passchier and B F M Bosnajakovic (Amsterdam: Elsevier) pp 83-7
MacLaughlin J A and Holick M F 1985 Ageing decreases the capacity of human skin to produce vitamin D3 J. Clin. Invest. 76 1536-8
Miescher G 1930 Das problem des Lichtschutzes und der Lichtgewohnung Strahlentherapie 35 403-43
Miller G T 1982 Living in the Environment 3rd edn (London: Wadsworth) pp 154-80
Molina M J and Rowland F S 1974 Stratospheric sink for chlorofluoromethanes: chlorine atom-catalyzed destruction of ozone Nature 249 810-2
Morison W L 1985 What is the function of melanin Arch. Dermatol. 121 1160-3
Moseley H, Davison M and MacKie R M 1983 Measurement of daylight UVA in Glasgow Phys. Med. Biol. 28 589-97
Muir C, Waterhouse J, Mack T, Powell J and Whelan S Eds 1987 Cancer Incidence in five Continents vol 5 IARC Scientific Publications 88 (Lyons: IARC)
NAS 1979 Protection Against Depletion of Stratospheric Ozone by Chlorofluorocarbons (Washington, DC: National Academy Press)
-----1984 Causes and Effects of Changes in Stratospheric Ozone: Update 1983 (Washington DC: National Academy Press)
Olson R L, Sayre R M and Everett M A 1966 Effect of anatomic location and time on ultraviolet erythema Arch. Dermatol. 93 211-5
OPCS 1981 Cancer Statistics: Incidence Survival and Mortality in England and Wales (London: HMSO) p 62
Paltridge G W and Barton I J 1978 Erythemal ultraviolet radiation distribution over Australia--the calculations, detailed results and input data Division of Atmospheric Physics Technical Paper 33 (Australia: Commonwealth Scientific and Industrial Research Organisation)
Parrish J A, Jaenicke K F and Anderson R R 1982 Erythema and melanogenesis action spectra of normal human skin Photochem. Photobiol. 36 187-91
Pathak M A and Faneslow D L 1983 Photobiology of melanin pigmentation: dose/response of skin to sunlight and its contents J. Am. Acad. Dermatol. 9 724-33
Patrick M H and Rahn R O 1976 Photochemistry of DNA and polynucleotides: photoproducts Photochemistry and Photobiology of Nucleic Acids vol II ed S Y Wang (New York: Academic) pp 35-95
Peak M J and Peak J G 1986 Molecular photobiology of UVA. Biological Effects of UVA Radiation ed F Urbach and R W Gange (New York: Praeger) pp 42-52
Pitts D G, Cullen A P and Parr W H 1976 Ocular Ultraviolet Effects from 295 nm to 335 nm in the Rabbit Eye (Washington, DC: US Department of Health, Education, Welfare (NIOSH))
Pitts D G and Tredici T J 1971 The effects of ultraviolet on the eye Am. Ind. Hyg. Assoc. J. 32 235-46
Preece M A, Tomlinson S, Ribot C A, Pietrek J, Korn H T, Davies D M, Ford J A, Dunnigan M G and O'Riordan J L H 1975 Studies of vitamin D deficiency in man Q. J. Med 44 575-89
Rigel D S, Kopf A W and Friedman R J 1987 The rate of malignant melanoma in the United States: are we making an impact? J. Am. Acad. Dermatol. 17 1050-2
Robertson D F 1972 Solar ultraviolet radiation in relation to human sunburn and skin cancer PhD Thesis University of Queensland
Rosen C F, Jacques S L, Stuart M E and Gange R W 1990 Immediate pigment darkening: visual and reflectance spectrophotometric analysis of action spectrum Photochem. Photobiol. 51 583-8
Rosen E S 1986 Pseudophakia and hazards of non-ionising radiation Semin. Opthalmol. 1 68-79
Roy C R, Gies H P and Elliott G 1989 The ARL solar ultraviolet radiation measurement programme Trans. Menzies Foundation 15 71-6
Rundel R 1986 Computation of spectral distribution and intensity of solar UV-B radiation Stratospheric Ozone Reduction, Solar Ultraviolet Radiation and Plant Life ed R C Worrest and M C Caldwell pp 49-62, 361-4 (Berlin: Springer)
Rupert C S 1982 Photobiological dosimetry of environmental ultraviolet radiation The Role of Solar Ultraviolet Radiation in Marine Ecosystems J Calkins (New York: Plenum) pp 131-41
Schothorst A A, Slaper H, Schouten R and Suurmond D 1985 UVB dose in maintenance psoriasis phototherapy versus solar UVB exposure Photodermatol. 2 213-20
Schothorst A, Slaper H, Telgt D, Alhadi B and Suurmond D 1987 Amounts of ultraviolet (UVB) received from sunlight or artificial UV sources by various population groups in the Netherlands Human Exposure to Ultraviolet Radiation: Risks and Regulations ed W F Passchier and B F M Bosnjakovic (Amsterdam: Elsevier) pp 269-73
Scotto, J, Cotton G, Urbach F, Berger D and Fears T 1988 Biologically effective ultraviolet radiation: surface measurements in the United States 1974-1985 Science 239 762-4
Scotto J, Fears T R and Fraumeni J F 1981 Incidence of Non-Melanama Skin Cancer in the United States (Washington, DC: US Department of Health and Human Sciences (NIH))
Setlow R B 1974 The wavelengths in sunlight effective in producing skin cancer: a theoretical analysis Proc. Nat. Acad. Sci. USA 71 3363-6
Slaper H and van der Leun J C 1987 Human exposure to ultraviolet radiation: quantitative modelling of skin cancer incidence Human Exposure to Ultraviolet Radiation: Risks and Regulations ed W F Passchier and B F M Bosnjakovic (Amsterdam: Elsevier) pp 155-71
Sliney D H 1987 Estimating the solar ultraviolet radiation exposure to an intraocular lens implant J. Cataract Refract Surg. 13 296-301
Smith G J 1990 A solar erythemal radiation monitoring programme in New Zealand and Queensland, Australia NZ Med. J. 103 5-6
Smith R C 1989 Ozone, middle ultraviolet radiation and the aquatic environment Photochem. Photobiol. 50 459-68
Smith R C and Baker K S 1979 Penetration of UV-B and biologically effective dose-rates in natural waters Photochem. Photobiol. 29 311-23
Smith R C, Baker K S, Holm-Hanson O and Olson R 1980 Photoinhibition of photosynthesis in natural waters Photochem. Photobiol. 31 585-92
Sober A J 1987 Solar exposure in the etiology of cutaneous melanoma Photodermatology 4 23-31
SORG 1990 United Kingdom Stratospheric Ozone Review Group Third Report (London: HMSO)
Stamp T C B and Round J M 1974 Seasonal changes in human plasma levels of 25-hydroxyvitamin D Nature 247 563-5
Sterenborg H J C M and van der Leun J C 1987 Action spectra for tumorigenesis by ultraviolet radiation Human Exposure to Ultraviolet Radiation: Risks and Regulations W F Passchier and B F M Bosnjakovic (Amsterdam: Elsevier) pp 173-90
Stern R S, Weinstein M C and Baker S G 1986 Risk reduction for non-melanoma skin cancer with childhood sunscreen use Arch Dermatol. 122 537-45
Swan C H J and Cooke W T 1971 Nutritional osteomalacia in immigrants in an urban community Lancet ii 456
Swanbeck G and Hillstrom L 1971 Analysis of etiological factors of squamous cell skin cancer of different locations Acta. Dermatol. Venereol., Stockholm 51 151-6
Tarrant A W S 1975 Further studies of the spectral power distribution of daylight in the ultraviolet region Compte Rendus 18th Session CIE, Publication 36 (London: CIE) pp 384-92
Taylor H R 1989 The biological effects of UV-B on the eye Photochem. Photobiol. 50 489-92
Taylor H R, West S K, Rosenthal F S, Munoz B, Newland H S, Abbey H and Effectt E A 1988 Effect of ultraviolet radiation on cataract formation New Engl. J. Med. 319 1429-33
Taylor H R, West S K, Rosenthal F S, Munoz B, Newland H S and Emmett E A 1989 Corneal changes associated with chronic ultraviolet irradiation Arch. Opthalmol. 107 1481-4
Teramura A H 1983 Effects of ultraviolet-B radiation on the growth and yield of crop plants Physiol. Plant. 58 415-27
-----1986 Overview of our current state of knowledge of UV effects on plants Effects of Changes in Stratospheric Ozone and Global Climate Vol I ed J G Titus (Washington, DC: US Environmental Protection Agency and United Nations Environmental Program) pp 165-73
Tevini M and Teramura A H 1989 UV-B effects on terrestrial plants Photochem. Photobiol. 50 479-87
Tucker M A, Shields J A, Hartge P, Augsburger J, Hoover R N and Fraumeni J F 1985 Sunlight exposure as risk factor for intraocular malignant melanoma New Engl. J. Med. 313 789-92
Urbach F 1969 Geographic pathology of skin cancer The Biologic Effects of Ultraviolet Radiation with Emphasis on the Skin ed F Urbach (Oxford: Pergamon) pp 634-50
-----1982 Photocarcinogenesis The Science of Photomedicine ed J D Regan and J A Parrish (New York: Plenum) pp 261-92
-----1987 Man and ultraviolet radiation Human Exposure to Ultraviolet Radiation: Risks and Regulations ed W F Passchier and B F M Bosnjaokovic (Amsterdam: Elsevier) pp 3-17
Urbach F, Epstein J H and Forbes P D 1974 Ultraviolet carcinogenesis: experimental, global and genetic aspects Sunlight and Man: Normal and Abnormal Photobiological Responses ed M A Pathak, L C Harber, M Seiji and A Kukita (Tokyo: University of Tokyo Press) 259-83
Vågerö D, Swerdlow A J and Beral V 1990 Occupation and malignant melanoma: a study based on cancer registration data in England and Wales and in Sweden Br. J. Indust. Med. 47 317-24
van der Leun J C 1984 UV-carcinogenesis Photochem. Photobiol. 39 861-8
Webb A R, DeCosta B R and Holick M F 1989 Sunlight regulates the cutaneous production of vitamin D3 by causing its photodegradation J. Clin. Endocrinol. Metab. 68 882-7
Webb A R and Holick M F 1988 The role of sunlight in the cutaneous production of vitamin D3 Ann. Rev. Nutr 8 375-99
Webb A R, Kline L W and Holick M F 1988 Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin J. Clin. Endocrinol. Metab. 67 337-8
Weiss J S, Ellis C N, Headington J T and Voor Lees J J 1988 Topical tretinoin in the treatment of ageing skin. J. Am. Acad Dermatol. 19 169-75
West S K, Rosenthal F S, Bressler N M, Bressler S B, Munoz B, Fine S L and Taylor H R 1989 Exposure to sunlight and other risk factors for age related macular degeneration Arch. Ophthalmol. 107 875-9
Wester U 1984 Solar ultraviolet radiation in Stockholm--examples of spectral measurements and influences of measurement error parameters Department of Radiation Physics, Karolinska Institute Internal Report RI 1984-03, Sweden
-----1987 Solar ultraviolet radiation on the Canary Islands and in Sweden--a comparison of irradiance levels Human Exposure to Ultraviolet Radiation: Risks and Regulations ed W F Passchier and B F M Bosnjakovic (Amsterdam: Elsevier) pp 275-9
Wetzler V R 1959 Zum Wirkungsspektrum der kancerogenen Eigenschaften des ultravioletten Lichtes Arch. Geschwuelstforsch. 14 120-131, 15 227-37
Wheldon T E 1988 Mathematical Models in Cancer Research (Bristol: Adam Hilger) pp 134-55
Whittaker R H and Likens G E 1975 The biosphere and man Primary Productivity of the Biosphere Ecological studies 14 ed H Lieth and R H Whittaker (Berlin: Springer-Verlag) pp 305-28
Worrest R C 1986 The effect of solar UV-B radiation on aquatic systems: an overview Effects of Changes in Stratospheric Ozone and Global Climate vol I ed J G Titus (Washington, DC: US Environmental Protection Agency and United Nations Environmental Program) pp 175-91
Young A R 1990 Senescence and sunscreens Br. J. Dermatol. 122 (Suppl. 35) 111-4
Young A R, Harrison J A and Walker S L 1988 Low SPF sunscreens reduce photoageing in mice Cosmet Toiletries 103 49-51
Zigman S T, Yuol T and Schultz J 1974 Cataract induction in mice exposed to near UV light Opthalmic Res. 6 259-70
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
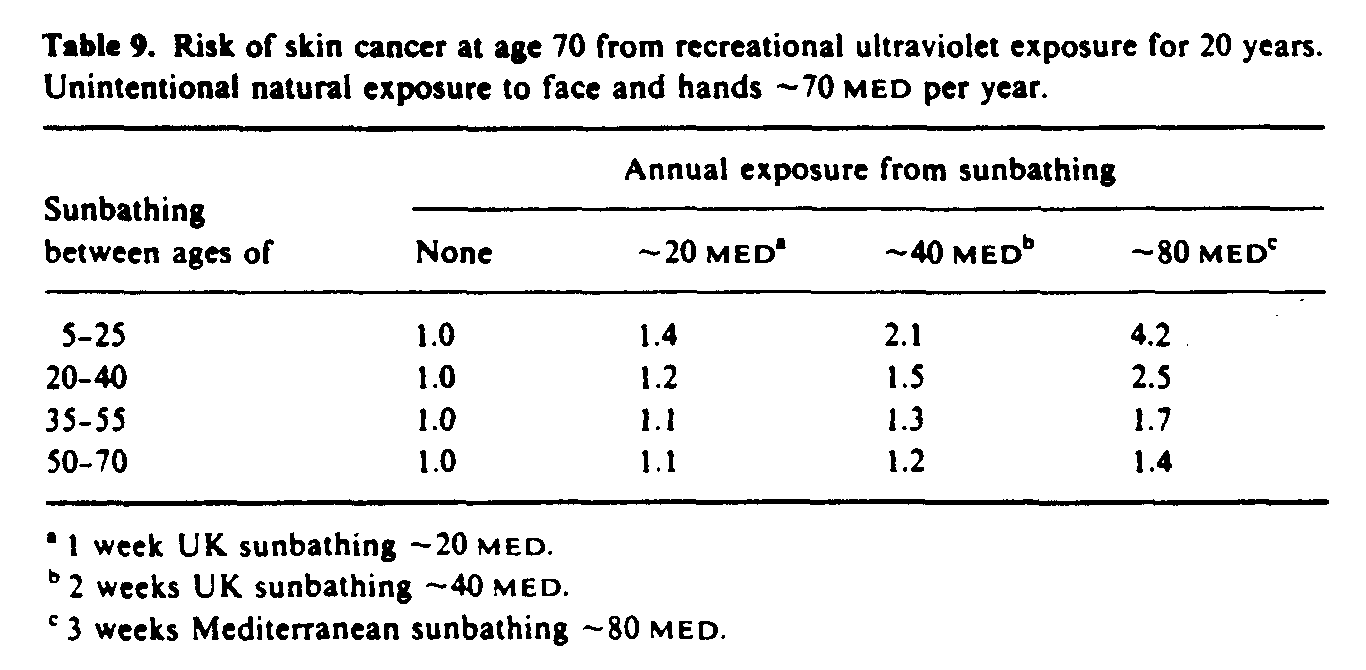{kind=link}
{kind=link}