 Reproduced, with permission, from:
Giannini, S. H. 1986. Effects of UVB on infectious diseases. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 101-12. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Reproduced, with permission, from:
Giannini, S. H. 1986. Effects of UVB on infectious diseases. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 101-12. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Volume 2: Stratospheric Ozone
Edited by James G. Titus U.S. Environmental Protection Agency
Suzanne Holmes Giannini
Columbia University College of Physicians and Surgeons New York, New York USA
The skin is our interface with the environment and our bodies' first line of defense against infectious agents. Other papers in this volume illustrate very elegantly that UV-B irradiation has a selective, suppressive effect on immune system functioning. They show us, further, that the skin itself is an immune organ, in that certain types of lymphoid cells are found predominantly in skin (Daynes et al. this volume; De Fabo and Noonan this volume; Elmets et al. this volume). The function of this skin-associated lymphoid tissue is to respond to antigens that enter the body by way of the skin. This cutaneous immune surveillance system is vital to our ability to resist invasions by infectious agents.
Infectious diseases remain serious public health problems in the tropics and semitropics. These are the geographic areas that also receive the highest levels of solar UV-B radiation in the world (Schulze 1970). A critical question arises: What are the effects of UV-B irradiation on the body's response to infectious agents penetrating the skin?
There are few experimental data to answer this important question. Analysis is further complicated because infection and disease outcome are affected, not only by the immunogenetic background of the host, but also by other environmental factors such as humidity, temperature, vector density and efficiency, concomitant infections, and host nutrition, to name but a few. Although the issue is very complex, a growing body of evidence suggests that UV-B irradiation can affect both pathogenesis and immunity in infectious diseases involving skin.
The effect of ultraviolet radiation on the progression of skin disease has been the subject of investigation for almost a century. In the late 1880s, the Danish physician Niels Finsen began a study of the interaction of skin with sunlight. By a series of experiments in which various wavelengths were filtered out from sunlight, he concluded that it was the ultraviolet component, which he called the "chemical rays," that caused inflammation and sunburn. In 1894, Finsen reported that the "chemical rays" of sunlight promoted scarring in smallpox, and that patients sequestered in rooms thickly hung with red curtains healed their pustules without scarring (Finsen 1901a). This illustrated that solar radiation could affect the outcome of infectious disease, and launched him on a study of ultraviolet radiation in the treatment of tuberculosis (Finsen 1901b). In 1903, Finsen received the Nobel Prize in Medicine for successful treatment of lupus vulgaris (tuberculosis of the skin) by local application of ultraviolet irradiation. Over the next four decades, ultraviolet light applied locally to skin was used to treat a variety of infectious diseases, including erysipelas, which had a 10% fatality rate until UV therapy appeared (Licht 1983). The advent of sulfones and antibiotics supplanted UV therapy, which has virtually disappeared from the infectious disease armamentarium.
As Finsen astutely observed after his discovery of the promotion of sunburn by UV, "What is more natural than that chemical rays should exert an injurious influence upon a diseased skin, when we see such severe inflammation produced by their influence upon the healthy skin?" (Finsen 1901a). The infectious diseases that are likely to be affected by UV are of two types.
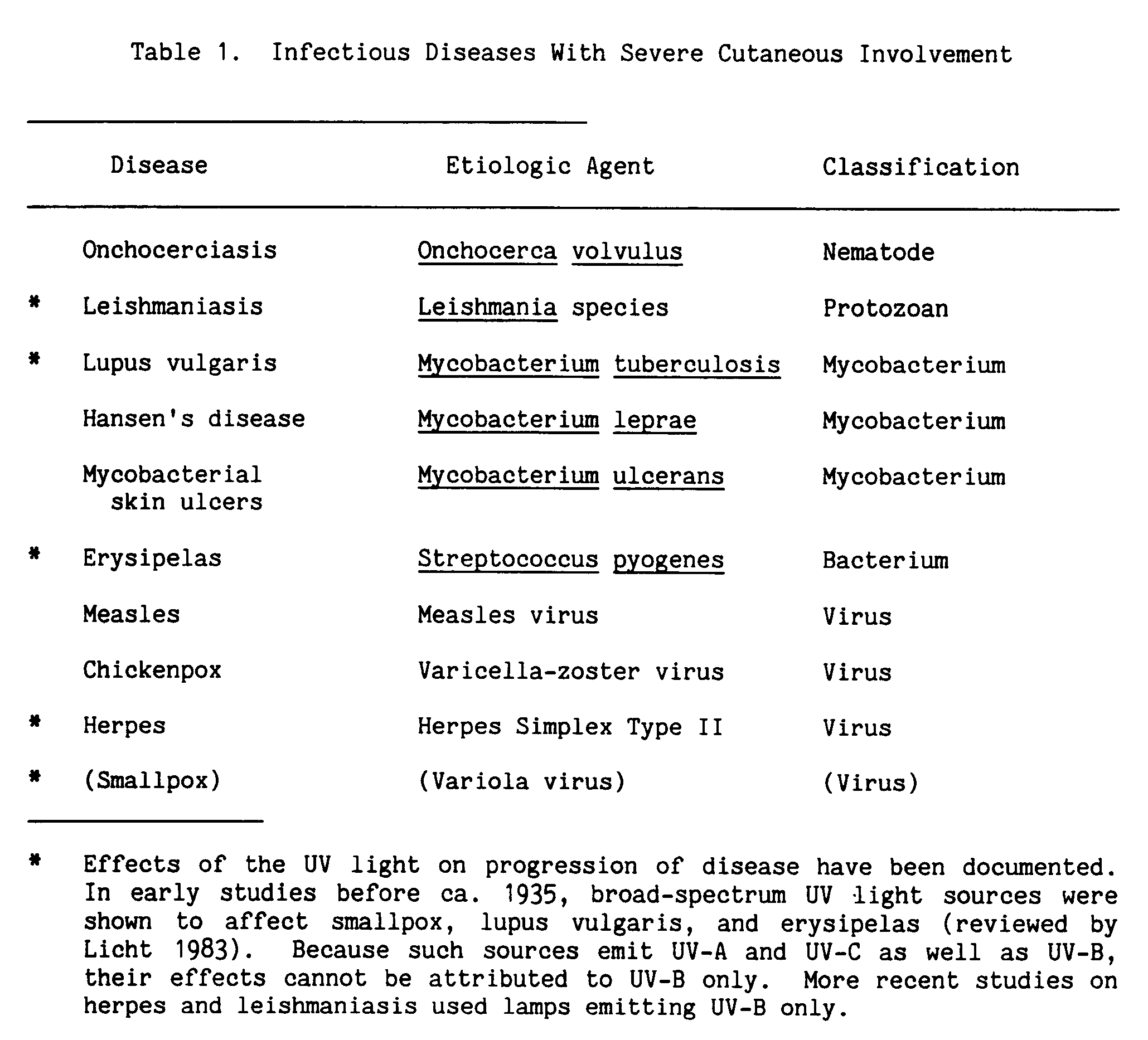
Infections that have severe cutaneous pathology, damaging the skin, are listed in Table 1. The effect of ultraviolet light on the progression of disease has been documented for five of these.
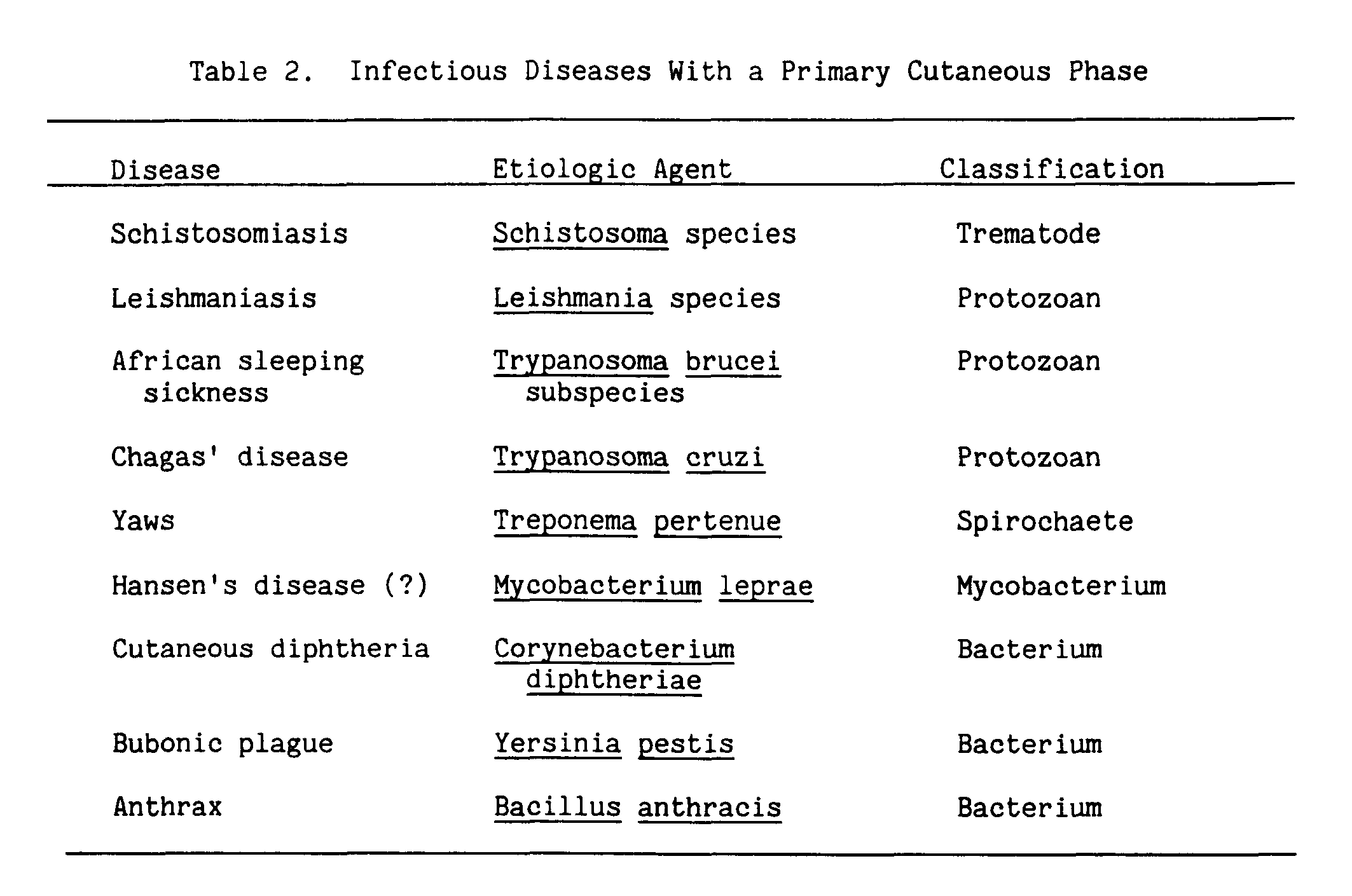
But perhaps of more concern to us here are those infectious agents for which the skin-associated lymphoid tissue is the first encounter with the host's immune system (Table 2). These diseases have a primary cutaneous phase, often asymptomatic, during which protective immunity to later infections can develop. Such diseases are of particular concern, because antigens that reach the immune system via skin that has been UV-B irradiated not only escape the immune surveillance system of the skin, but also can make the host tolerant to later exposure to the same antigens (Kripke 1984; Elmets et al. 1983). For most of the diseases with a primary cutaneous phase, the development of cell-mediated immune responses is necessary for protection. Other papers in this volume show that cell-mediated immunity is more critically affected by UV-B.
As a model for the effects of UV-B on infectious disease, let us turn to leishmaniasis in the mouse. Leishmania have an initial cutaneous phase, which can be asymptomatic and precede more serious sequelae; and they also cause skin ulcers (Tables 1 and 2).
The leishmaniases are a spectrum of diseases, caused by members of the genus Leishmania, which are obligate intracellular protozoan parasites living with macrophages and monocytes. Paradoxically, for Leishmania to survive in the body, they must be ingested by macrophages, whose function normally is to digest invading microorganisms. The parasites are transmitted from host to host by phlebotomine sandflies, which deposit the Leishmania in the upper layer of the dermis or in the epidermis. Phlebotomine proboscides are too short to penetrate protective clothing, so that sandflies must feed on exposed skin.
Most infections with Leishmania species lead to self-healing skin ulcers, which leave the patient permanently scarred but immune to reinfection with related species of Leishmania. However, more serious sequelae are noted in some forms of leishmaniasis. In chiclero's ulcer, the entire ear pinna can be eroded away; in mucocutaneous leishmaniasis, parasites metastasize from the original skin lesion to the mucocutaneous junctions. These severe mutilating lesions do not heal spontaneously, and they occur on parts of the body that receive high amounts of UV-B in sunlight (Urbach 1969). A primary skin ulcer does not always precede the onset of the most fatal form of leishmaniasis, kala-azar, in which the Leishmania invade the entire reticuloendothelial system. Kala-azar smoulders for years in endemic foci, to erupt into fulminating epidemics in times of migration, famine, and war.
Leishmanial disease occurs on every continent except Australia and Antarctica. There are at least four different species causing human disease in the Old World, and at least three in the New World. All Leishmania species can cause skin ulcers, but only some of them can cause more severe disease in immunogenetically predisposed individuals. Yet because not all infected persons in groups at high risk for serious sequelae develop severe disease, other factors are probably involved. In countries endemic for leishmaniasis, skin is naturally exposed both to the bites of infective sandflies and to UV-B in sunlight, which is known to depress the functions of the skin-associated lymphoid tissue. So it seemed likely that early immunological events occurring in the skin could critically determine the outcome of infection with Leishmania.
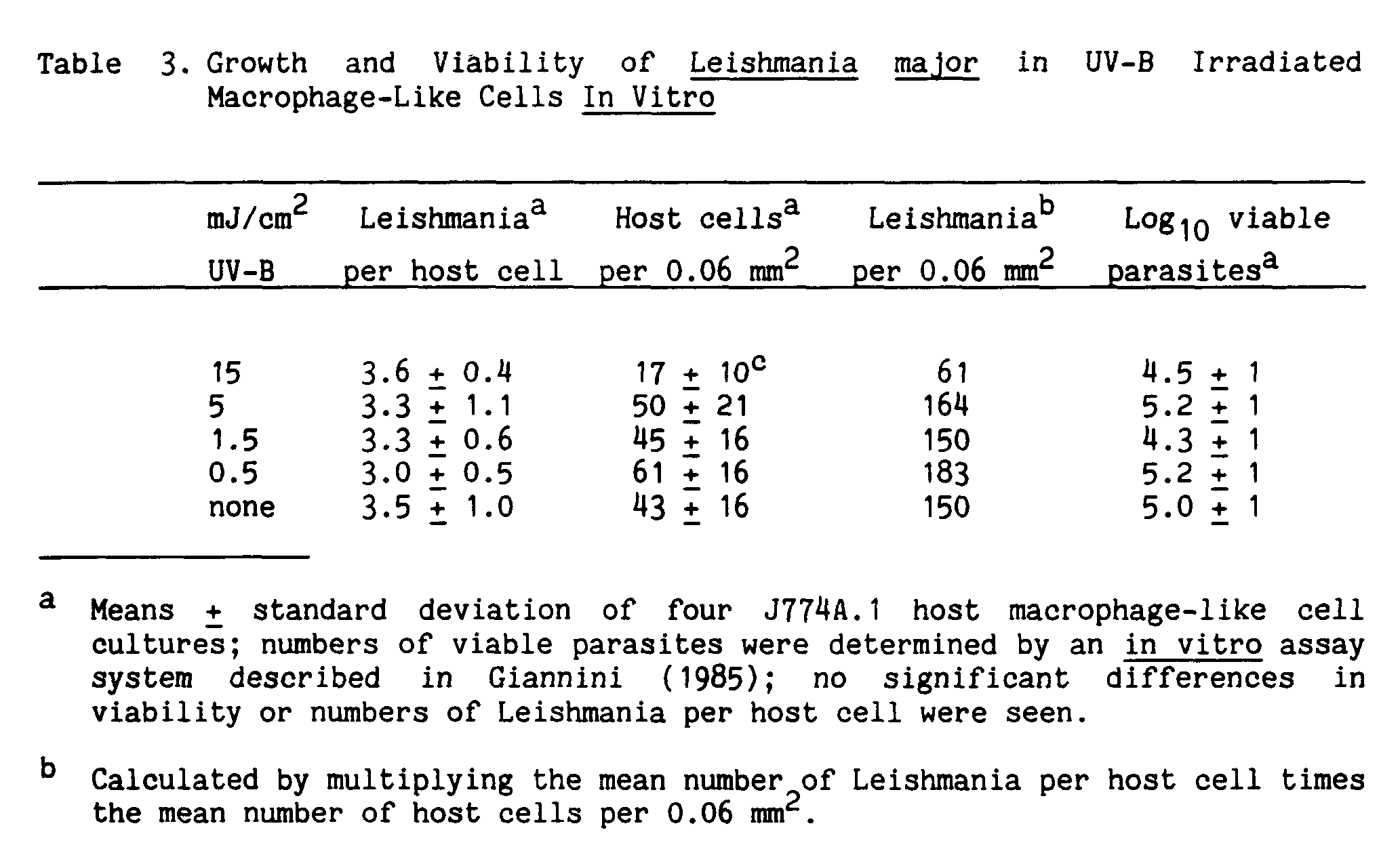
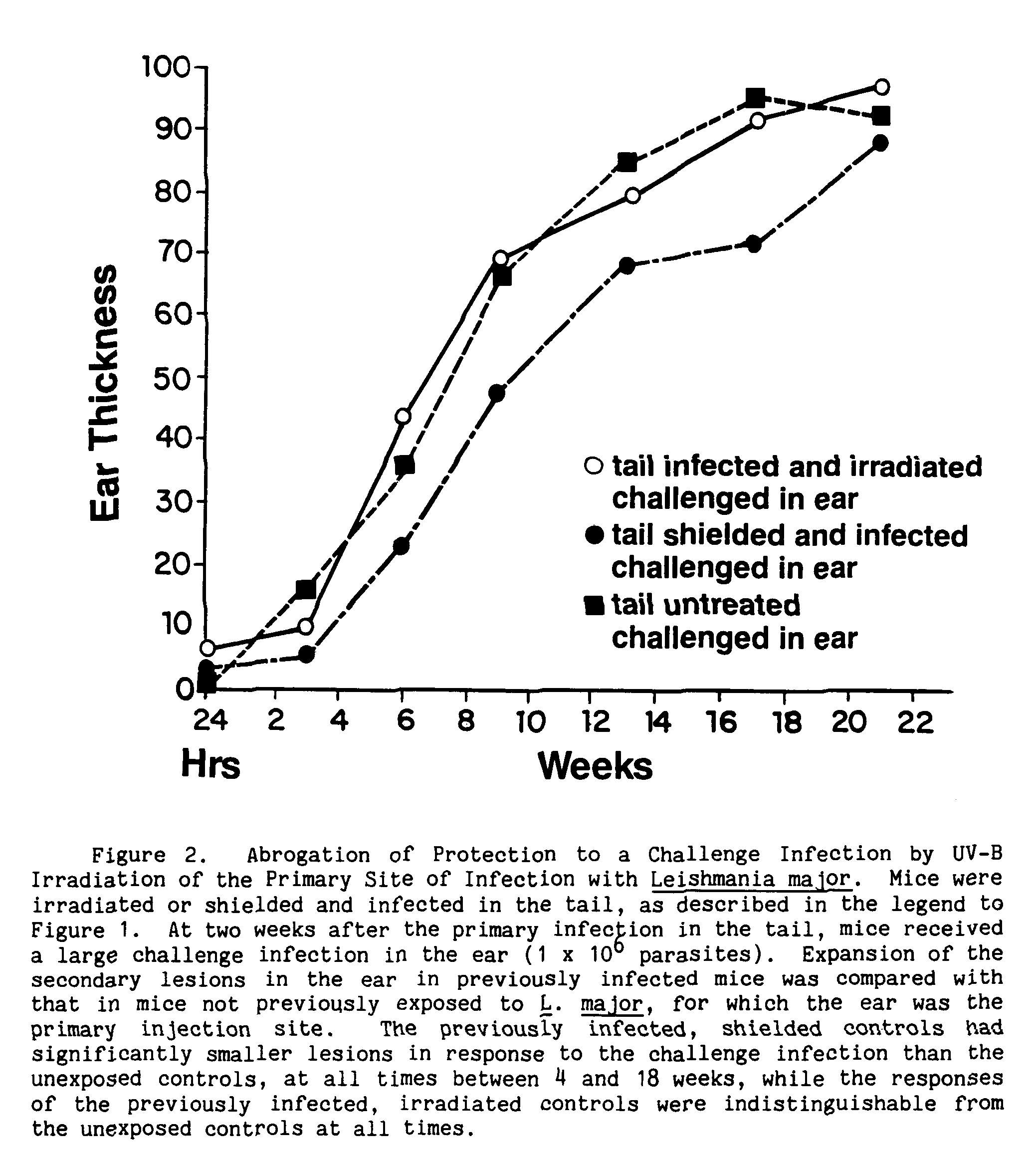
In a series of experiments designed to evaluate the effects of UV-B on the development of cutaneous leishmaniasis in mice (Giannini 1986), I found that local irradiation of the injection site with low doses of UV-B around the time of initial infection indeed affected lesion development (Figure 1). The doses used were not high enough to cause sunburn, but were about the amount of UV-B in an hour's exposure to sunlight in the semitropics on a bright day at noon. Surprisingly, lesion severity was reduced in the irradiated mice (Giannini 1986). The main targets of the UV-B were host cells, and not Leishmania, because the parasites were present in the skin and in vitro in the same numbers in both irradiated and control groups (Tables 3 and 4). But despite their apparently healthy skin, the irradiated mice had parasites at the injection site and in their local lymph nodes (Table 5). When they were tested for their ability to mount cell-mediated immune responses to leishmanial antigens, the UV-B irradiated mice had almost no immunity, compared with the unirradiated control mice (Table 6). Their lack of cell-mediated immunity reduced the irradiated mice's ability to control a second challenge infection in a skin site different from that of their first exposure, which had been irradiated (Figure 2). Since Leishmania may persist in the local lymph nodes, long after skin lesions have healed, and even when parasites cannot be detected in the skin (Hill, North, and Collins 1984; Titus et al. 1985; Giannini, unpublished), it is possible that depression of the host immune response by other factors such as stress or impaired nutrition could put the infected, though asymptomatic, individual at risk for disseminated disease from the parasites lingering in the lymph nodes.
A similar suppressive effect of UV-B on cell-mediated immunity to herpes simplex Type II virus has also been shown (Hayashi and Aurelian 1986). Exposure to UV-B during the primary phase of infection suppresses the development of cell-mediated immune responses to the virus (Howie, Norval, and Maingay 1986), and irradiation of healed skin lesions triggers recurrent herpes infections (Wheeler 1975; Blyth et al. 1976).
From these and other data we can conclude that local perturbations in the functions of the skin-associated lymphoid tissue during cutaneous infections can profoundly influence the immunological response to antigens of invading microbes and the subsequent development of clinical disease.
If the immune functions of the skin cannot be protected from the effects of UV-B by the use of sunscreens (Menter this volume), then it may be difficult to avoid exposure to UV-B, especially in agricultural societies in the tropics and semitropics, where heat makes protective clothing impractical to wear.
Increases in the amount of solar UV-B reaching the earth's surface will likely affect the severity of infectious disease. More precise predictions are not possible, because wavelengths and dosages will critically affect the response curve. In addition, the global effects of UV-B include other environmental factors that impact on disease development, such as climate, vector density, and food availability. It is probable that some cutaneous infectious diseases will be exacerbated by increased UV-B radiation, as is the case with smallpox and herpes. In other cases, pathogenesis of cutaneous disease may be suppressed, such as in cutaneous leishmaniasis and lupus vulgaris.
But we do well to remember that escape from overt skin disease may have a high price, namely, forfeiture of protective immunity to microbes that escape the immune surveillance network in the skin while its functions are inactivated by UV-B.
This work was supported by Public Health Service Grant AI 18937 from the National Institutes of Health. I thank Dr. Edmond A. Goidl for many helpful discussions, and Ms. Susie Mathews for expert technical assistance.
Blyth, W.A., T.J. Hill, H.J. Field, and D.A. Harbour. 1976. Reactivation of herpes simplex virus infection by ultraviolet light and possible involvement of prostaglandins. J. Gen. Virol. 33:547-50.
De Fabo, E., and F. Noonan. 1986. Urocanic acid and its role in the immune response. This symposium.
Elmets, C.A., P.R. Bergstresser, R.E. Tigelaar, and J.W. Streilein. 1983. In vivo low dose UV-B irradiation induces suppressor cells to contact sensitizing agents. In The effect of ultraviolet radiation on the immune system, ed. J.A. Parrish, 317-33. Skillman, New Jersey: Johnson & Johnson Baby Products Company.
Finsen, N.R. 1901a. The chemical rays of light and smallpox. In Phototherapy, 1-36. London: Edward Arnold.
Finsen, N.R. 1901b. The treatment of lupus vulgaris by concentrated chemical rays. In Phototherapy, 73-75. London: Edward Arnold.
Giannini, M.S.H. 1986. Suppression of pathogenesis in cutaneous leishmaniasis by UV irradiation. Infect. Immun. 51:838-43.
Giannini, M.S.H. Cutaneous leishmaniasis: A lymphatic infection. Manuscript submitted.
Giannini, S.H. 1985. Induction and detection of leishmanial infections in Rattus norvegicus. Trans. Roy. Soc. Trop. Med. Hyg. 79:458-61.
Hayashi, Y., and L. Aurelian. 1986. Immunity to herpes simplex virus type 2: Viral antigen-presenting capacity of epidermal cells and its impairment by ultraviolet irradiation. J. Immunol. 136:1087-92.
Hill, J.O., R.J. North, and F.M. Collins. 1984. Advantages of measuring changes in the number of viable parasites in murine models of experimental cutaneous leishmaniasis. Infect. Immun. 39:1087-94.
Howie, S., M. Norval, and J. Maingay. 1986. Exposure to low-dose ultraviolet radiation suppresses delayed-type hypersensitivity to herpes simplex virus in mice. J. Invest. Dermatol. 86:125-28.
Kripke, M.D. 1984. Immunological unresponsiveness induced by ultraviolet radiation. Immunolog. Rev. 80:87-102.
Licht, S. 1983. History of ultraviolet therapy. In Therapeutic Electricity and Ultraviolet Radiation, 3rd ed., ed. G.K. Stillwell, 174-93. Baltimore: Williams & Wilkins.
Schulze, R. 1970. Global radiation climate. Wiss. Forschungsber. 72:1-220.
Titus, R.G., M. Marchand, T. Boon, and J.A. Louis. 1985. A limiting dilution assay for quantifying Leishmania major in tissues of infected mice. Parasite Immunol. 7:545-55.
Urbach, F. 1969. Geographic pathology of skin cancer. In The biologic effects of ultraviolet radiation (with emphasis on the skin), ed. F. Urbach, 635-50. Oxford: Pergamon Press.
Wheeler, C.E., Jr. 1975. Pathogenesis of recurrent herpes simplex infections. J. Invest. Dermatol. 65:341-46.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}