 Reproduced, with permission, from:
Fisher, M. S., J. M Menter, L. Tiller, and I. Willis. 1986. Sunscreens do not abrogate UV-induced suppression of contact hypersensitivity. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 131-40. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Reproduced, with permission, from:
Fisher, M. S., J. M Menter, L. Tiller, and I. Willis. 1986. Sunscreens do not abrogate UV-induced suppression of contact hypersensitivity. In Stratospheric ozone. Vol. 2 of Effects of changes in stratospheric ozone and global climate, ed. J. G. Titus, 131-40. Proceedings of the United Nations Environment Programme (UNEP)/Environmental Protection Agency (EPA) International Conference on Health and Environmental Effects of Ozone Modification and Climate Change. Washington, D.C.: U.S. Environmental Protection Agency.
Edited by James G. Titus
M.S. Fisher, J.M. Menter, L. Tiller, and I. Willis Morehouse School of Medicine Atlanta, Georgia USA
Contact hypersensitivity (CHS) in mice can be induced by cutaneous sensitization followed by elicitation via ear-painting with trinitrochlorobenzene (TNCB). This CHS reaction is systemic and can be suppressed by exposure of mice to suberythemogenic doses of 280-315 nm radiation. In this paper, we investigate whether a commercially available water-resistant sunscreen (SPF-6) containing padimate 0 (UV-B absorber) and oxybenzone (UV-A absorber) was effective in preventing systemic suppression of CHS induced by either FS36 sunlamp exposure or solar simulating radiation. We observed that this sunscreen preparation could not prevent the immunologic suppression of contact hypersensitivity by UV radiation. These results indicate that application of sunscreen does not retard the development of suppression of CHS following repeated UV exposure under conditions where erythema is not clinically observed. Thus, erythema may not be a good end point for assessing systemic immune suppression and its consequences.
Exposure of mice to ultraviolet (UV) radiation systemically suppresses their ability to reject highly antigenic UV-induced skin cancers (Fisher 1977; Fisher and Kripke 1977). This state of unresponsiveness is mediated immunologically and is due in part to the presence of antigen-specific suppressor T lymphocytes (Fisher 1977, 1978; Fisher and Kripke 1977, 1978, 1982; Spellman and Daynes 1977). A second UV-induced immunosuppressive defect has also been described in which a single exposure of the dorsal skin of mice to UV radiation was sufficient to suppress contact hypersensitivity (CHS) to 2-chloro-1,3,5-trinitrobenzene (TNCB) applied to the abdominal surface (Noonan et al. 1981). The cellular basis for the suppression of contact hypersensitivity appears to involve a UV-induced alteration in the functional activity of antigen-presenting cells (Noonan et al. 1981; Greene et al. 1979).
The photobiologic and immunologic similarities of these two UV-induced systemic immunosuppressive events suggest that they may share common steps (Noonan et al. 1981; De Fabo and Kripke 1979; Noonan, De Fabo, and Kripke 1981a, b), which appear to be: (a) an alteration in the presentation of certain antigens, particularly those encountered via the cutaneous route (Noonan et al. 1981) and (b) the induction of T suppressor cells as a consequence of altered antigen presentation (Greene et al. 1979). The main difference between both forms of systemic immunologic suppression is that the antigen that induces the suppressor cells is applied exogenously in the case of contact hypersensitivity, but must be formed endogenously by UV exposure of the skin in the induction of suppressor T cells that regulate tumor rejection. Another important difference between the two systems is the actual dose of UV needed to induce 50% suppression. For CHS, the dose is approximately 13 times less than that needed to induce the tumor susceptibility in 50% of the UV-irradiated animals (Noonan, De Fabo, and Kripke 1981b).
Sunscreens, the majority of which use para-aminobenzoic acid (PABA) or one of its derivatives and/or a benzophenone derivative as their active ingredients, protect against a number of UV radiation effects. The ability of topically applied sunscreens to reduce erythema and skin damage caused by chronic UV exposure, expressed as a "sun protective factor" (SPF), is well documented (Pathak 1969; Willis and Kligman 1970; Kligman, Akin, and Kligman 1980, 1982). In addition, sunscreens have been shown to protect against both the cocarcinogenic as well as the carcinogenic effect of UV radiation (Snyder and May 1975; Kligman, Akin, and Kligman 1980; Wulf et al. 1982; Stern, Weinstein and Baker 1986). To date, however, very few studies have evaluated the effect of these sunscreen agents on the more subtle immunoregulatory effects now known to be produced by UV-B irradiation. One such study by Gurish et al. (1981) found that pretreatment of mice with a PABA sunscreen completely prevented the UVB-induced histologic changes in exposed skin and rendered the mice tumors susceptible, although this susceptibility could not be adoptively transferred to normal untreated animals by lymphoid cell injection. A later study (Morison 1984) with a similar sunscreen but with higher doses of UV-B radiation showed not only induction of the tumor-susceptible state, but also that this state could be transferred to syngeneic animals by lymphoid cells.
In this study, we report the results of experiments designed to evaluate the potential protective effect of a commercial sunscreen preparation on the immunologic suppression of contact hypersensitivity by UV radiation. We observed that this sunscreen preparation with PABA and oxybenzone as its active ingredients was incapable of preventing the immunologic suppression of contact hypersensitivity by UV radiation.
Animals
Inbred albino hairless mice which have been bred in our laboratories from the inbred HRA/Skh strain were obtained from Stanley Mann of Philadelphia. The hairless mice were from 8 to 16 weeks old at the start of an experiment, but in any one experiment, the age of the animals did not vary by more than 1 week. The animals had free access to Purina Lab Chow and water and were housed in rooms where ambient lighting was automatically regulated on a 12-hour light/dark cycle.
Contact Sensitization
Each experimental group utilized 5-10 hairless mice. Contact hypersensitivity was induced by the method of Asherson and Ptak (1968). The abdomen was painted with 100 l (microliters) of a 5% solution of trinitrochlorobenzene (TNCB) in acetone. The mice were tested for the development of contact hypersensitivity five days later by applying 5
l of a 1% TNCB in acetone to both surfaces of each ear. Ear thickness was measured with an engineer's micrometer (Model No. 7309, Mitutoyo, Japan) before and 24 hours after application of the challenge dose. The ear swelling obtained was compared to the ear swelling of control mice that were challenged, but not sensitized. The statistical significance of the differences in ear swelling between groups was evaluated using the Student's T-test.
Sunscreen Preparation
A commercial sunscreen (SPF-6) contained padimate O and oxybenzone as active ingredients. The sunscreen was applied uniformly to the dorsal skin of the mice at a final dose of 2 l/cm[2], 20 minutes prior to UV exposure.
Energy Sources and Measurements
Two energy sources were used during the experiments. One source was a 1.6 kW xenon arc solar-simulating lamp which has been previously described (Willis, Menter, and Whyte 1981). Energy output from this lamp system was measured by a calibrated Eppley Thermopile in conjunction with a Keithley millimicroyoltmeter (Model 149). The total radiation was determined to be 26.2 mW/cm[2], 86 cm (11.5 in) from the exit port. The second source was a bank of three FS36T12-UV-B-VHO lamps (Light Sources) housed in an Ultralite bench top irradiator [BT 3-36 VHO(UV-B)]. Energy output from this system was measured as above 19 cm (7.5 in) from the source and the total output was determined to be 1.4 mW/cm[2].
Minimal Erythema Dose (MED) Determination
For each UV energy source, five hairless mice were irradiated at 25% dose increments. Erythemal responses were noted 24 hours after irradiation. The MED was taken as the exposure required to elicit barely visible minimal redness. Each set of experiments was repeated three times. SPF values were assessed for a sunscreen preparation as the ratio of MED with sunscreen to MED without sunscreen.
Effect of UV Exposure on the Development of CHS
To assess the depression of CHS by UV treatment, mice were sensitized three days after the last UV treatment and challenged five days later. The net ear swelling (ear swelling of sensitized and challenged mice minus the ear swelling of unsensitized, but challenged mice) was determined for unirradiated and UV-treated mice. The percentage of control response was determined by the following formula: A-B/C-D X 100. The letters represent the ear swelling in: (A) mice exposed to UV and sensitized with TNCB; (B) mice exposed to UV, but not sensitized; (C) unirradiated and sensitized mice; and (D) unirradiated and unsensitized.
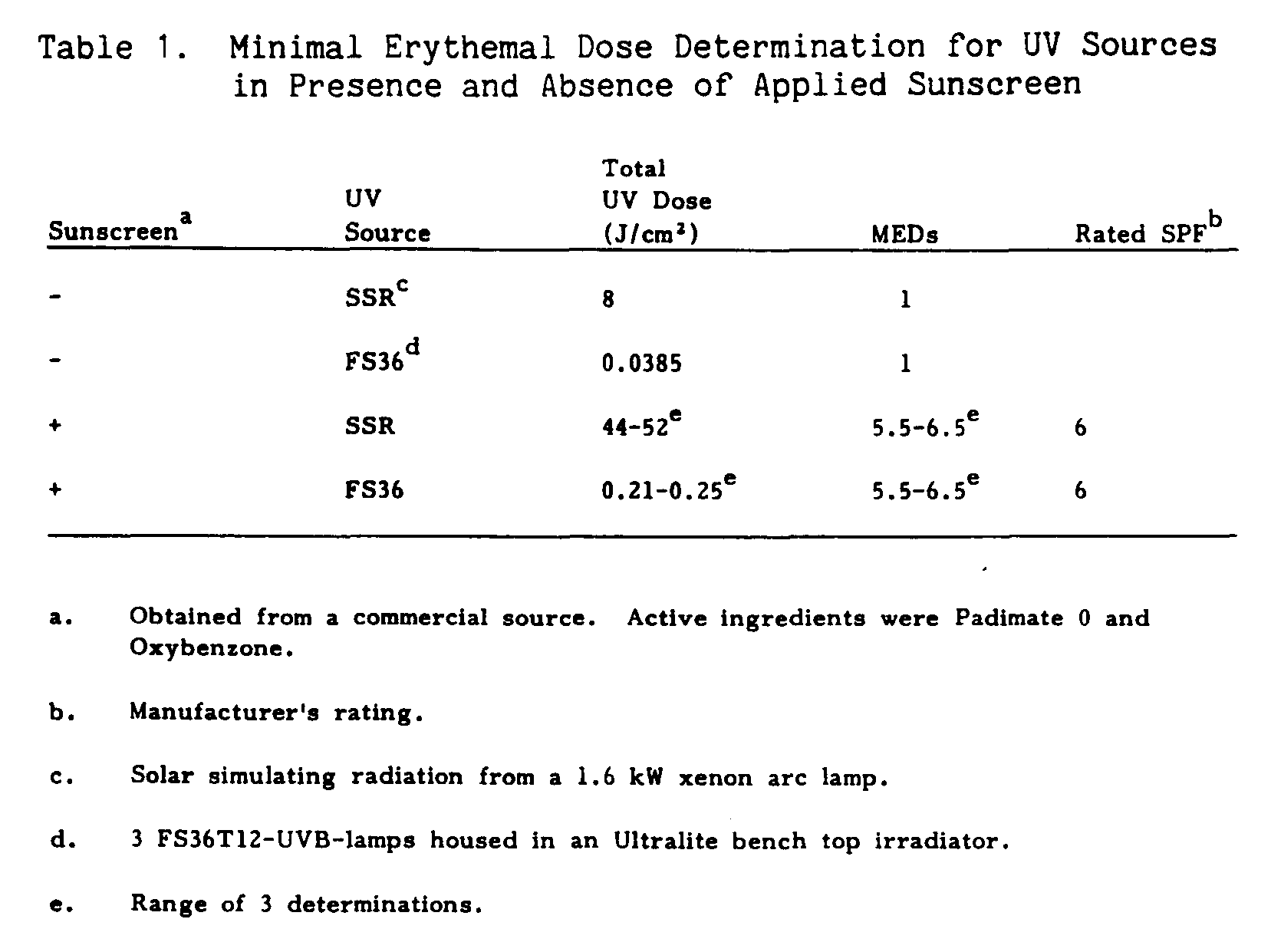
MED Determination
The doses required to elicit minimal erythema responses for solar-simulating radiation (SSR) and FS36 sunlamps are given in Table 1. Minimal erythemal dose determination for three FS36 sunlamps was determined to be 38.5 mJ/cm[2] while the total MED dose for SSR was 8 J/cm[2]. To test the MED of a commercially available sunscreen (SPF-6), sunscreen was applied to the dorsal surface of inbred hairless mice at a final dose of 2 l/cm[2], 20 minutes prior to UV exposure. For both radiation sources, the commercially available sunscreen was determined to have a MED of 6.0 +/- 0.5.
Topical Application of Sunscreen and UV-Induced Suppression of Contact Hypersensitivity
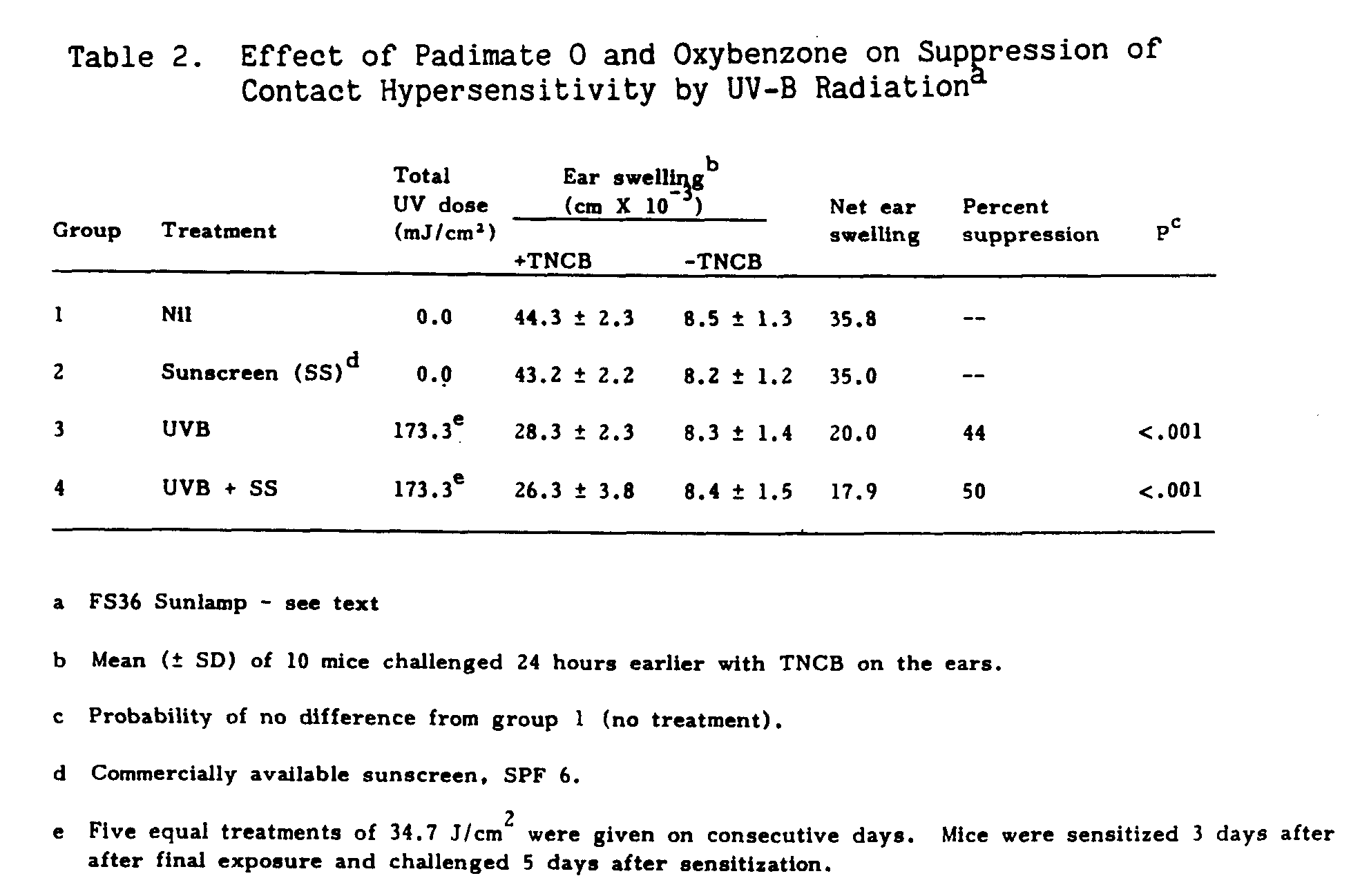
In the first set of experiments, two groups of ten mice were pretreated with sunscreen; one group served as an unirradiated control, group 2, while the other group, group 4, was irradiated with a bank of three FS36 sunlamps. During the treatment regimen, no erythematous response was observed in the group receiving the sunscreen preparation. Table 2 shows the results obtained from mice given sunscreen and treated with UV-B radiation. Sunscreen application had no effect on the contact hypersensitive response, as seen by the similarity in groups 1 and 2. The amount of suppression with (44%) and without (50%) topically applied sunscreen was similar.
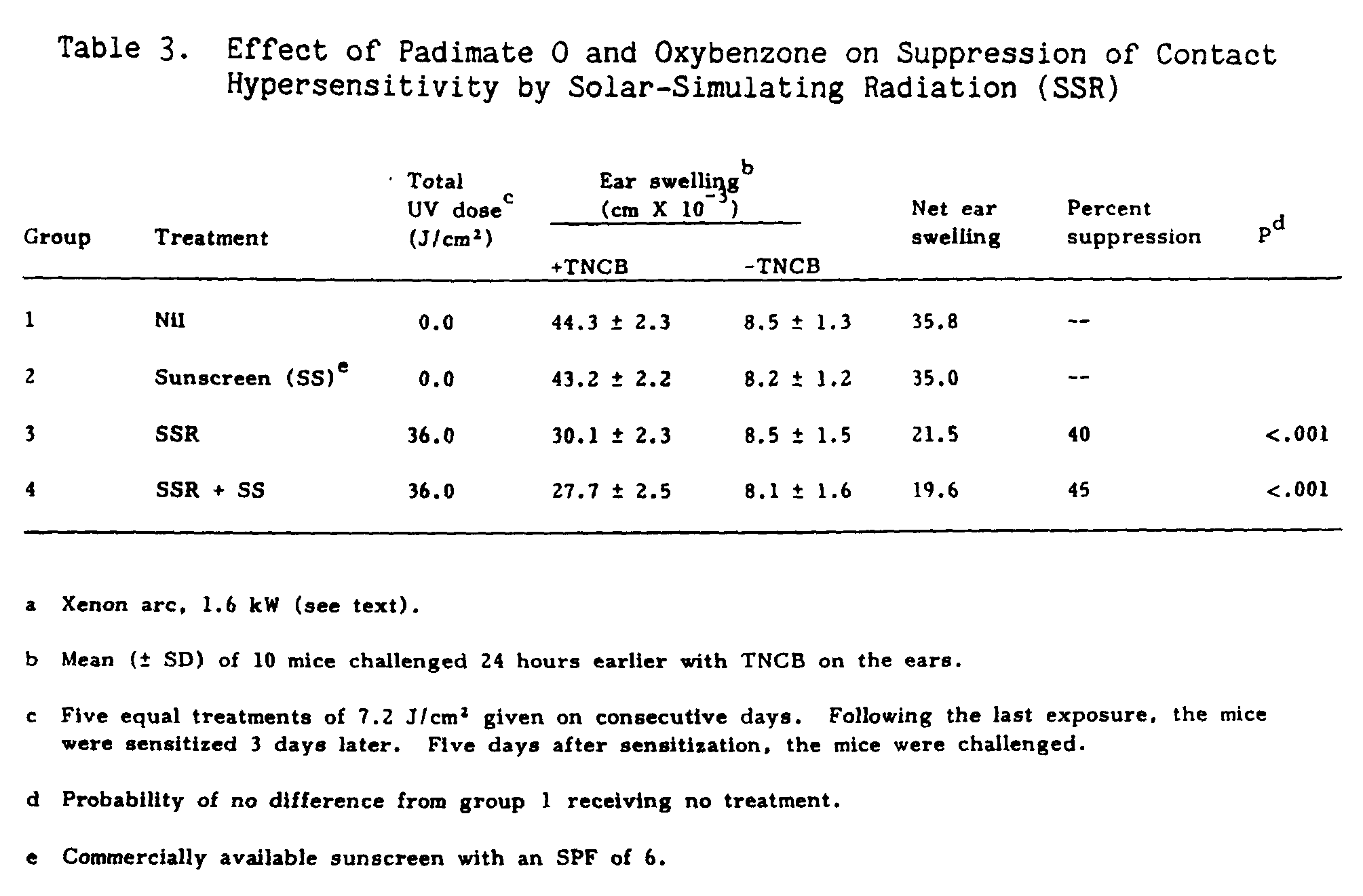
In the second part of the experiment (Table 3), the sunscreen preparation was topically applied to a group of ten mice receiving SSR radiation (group 4), and another group of ten mice, group 2, which served as a control. The results show that topical application of sunscreen had no effect on the ability of mice to mount a CHS response, groups 1 and 2. The presence of the sunscreen had no effect on suppression of contact hypersensitivity by SSR. The amount of suppression with applied sunscreen was 45% (group 4) while suppression from SSR alone (group 3) was 40%. As with the previous experiment, no erythematous response was observed in the group receiving the sunscreen preparation.
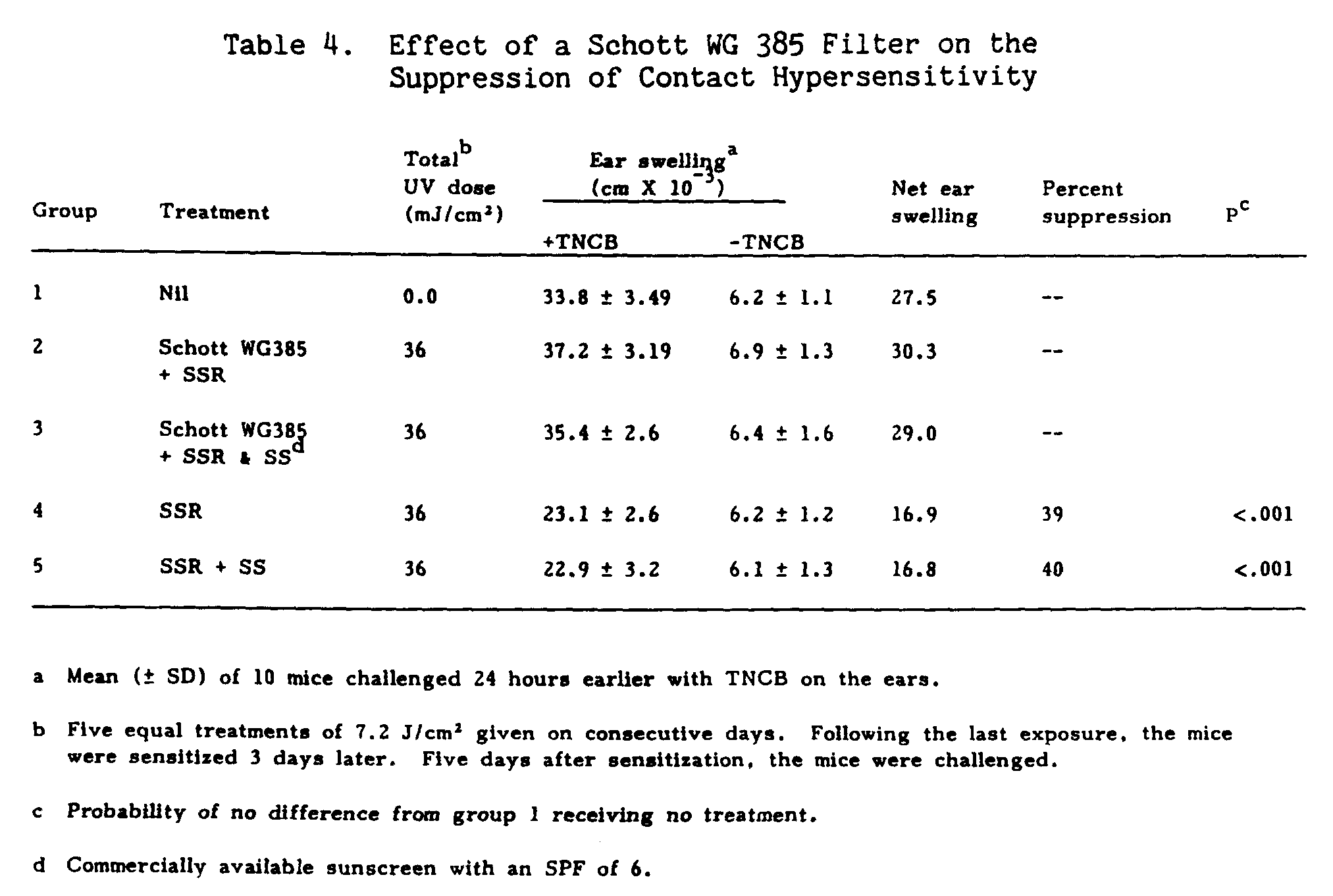
To determine if the previously observed suppression was due to heat alone, two groups of mice were exposed to SSR radiation through a Schott WG 385 filter, a cut off filter with 50% transmission at 385 nm (Table 4, groups 2 and 3). Group 3 received topical sunscreen application while group 2 was treated with SSR alone. The total UV dose of 36 J/cm[2] was unable to suppress the contact hypersensitivity response when filtered through a Schott WG 385 filter. However, in the absence of the Schott filter in both the SSR group alone (group 4) and the group receiving topical sunscreen plus SSR (group 5), suppression of contact hypersensitivity was observed.
Treatment of hairless mice with a commercial sunscreen (SPF-6) which contained padimate O and oxybenzone as the active ingredients did not prevent systemic immunologic suppression of contact hypersensitivity following exposure with either SSR or FS36 sunlamps. These results differ from those of Morison (1984), who found that pretreatment of C3H mice with PABA sunscreen (SPF-8) only induced 25% suppression of the contact hypersensitivity response following UV-B exposure, which he determined not to be significant. We used different mice, albino hairless, and a sunscreen with an SPF-6 rating. In addition, our total dose of UV was lower than his by a factor of 10. These differences alone could account for the discrepancy between Morison's and our results.
In Morison's study, the feet and nose were neither covered nor treated with sunscreen. Hence, the UV-induced suppression he observed in C3H mice could have been due to irradiated areas not treated with sunscreen. In our experiments, the head and neck were covered with a shield and the back, feet, and tail treated with sunscreen. Thus, our results are difficult to explain by exposure of non-sunscreen treated sites.
If mice are irradiated with polychromatic UV radiation, which was insufficient to elicit an erythema "sunburn" response, then UV-A rather than UV-B might be responsible for the induction of immune suppression observed in our experiments. However, because previous studies have shown that wavelengths shorter than 320 nm are responsible for systemic immunologic suppression (Noonan et al. 1981a; De Fabo and Kripke 1980), other alternatives are considered. The finding that the sunscreen did not abrogate the effects of UV radiation on contact hypersensitivity might be explained by assuming that sunscreen agents act as photosensitizers initiating a photochemical reaction in the skin that could then lead to the suppression of contact hypersensitivity (Gurish et al. 1981). In support of this theory, Hodges et al. (1977) found increased genetic damage when E. coli were irradiated in the presence of PABA. Another explanation is that the amount of energy not absorbed by the sunscreen agent is sufficient to induce the suppression of contact hypersensitivity. The location of the sunscreen in the skin may be very important in this regard. Willis and Kligman (1970) demonstrated that significant photoprotection from PABA persisted despite repeated stripping of the stratum corneum. These results suggest that sunscreens may "pool" beneath the stratus corneum. Recently, De Fabo and Noonan (1983) suggested that a photoreceptor, urocanic acid, resides close to the surface of the skin in the stratum corneum. This receptor mediates the initial event in systemic immunologic suppression by UV radiation. Our results are consistent with their interpretation: for if sunscreens pool beneath the stratum corneum, this would allow exposure of the photoreceptor, which is in the stratum corneum and above the sunscreen, so that subsequent events associated with systemic immunologic suppression could take place.
PABA absorbs radiation between 270 and 320 nm very efficiently, but not wavelengths below 270 nm. The experiment using the FS36 lamps could be explained by radiation exposure below 270 nm because no filter was used to cut out these wavelengths. However, the experiment that used SSR, which emits essentially no irradiation less than 290 nm, cannot be explained as due to UV-C radiation not absorbed by the sunscreen.
Because sunscreens are known to protect against both skin damage and the carcinogenic effects of UV radiation, our results seem to suggest a separation of UV-induced carcinogenic effects from the systemic immunologic effects. Our results also suggest that if these results can be generalized to humans, the use of sunscreens may not provide the protection needed to abrogate the immunologic effects of UV radiation. Thus, even sunscreen users would be at increased risk for skin cancer, because altered antigen presentation would already have been established and the formation of suppressor T cells would appear earlier in these individuals than in those persons who have never had any sun exposure.
Asherson, G.L., and W. Ptak. 1968. Contact and delayed hypersensitivity in the mouse. I. Active sensitization and passive transfer. Immunol. 15:405-16.
De Fabo, E.C., and M.L. Kripke. 1979. Dose-response characteristics of immunologic unresponsiveness to UV-induced tumors produced by UV-irradiation of mice. Photochem. Photobiol. 30:385-90.
De Fabo, E.C., and M.L. Kripke. 1980. Wavelength dependence and dose-rate independence of UV radiation-induced immunologic unresponsiveness of mice to a UV-induced fibrosarcoma. Photochem. Photobiol. 32:183-88.
De Fabo, E.C., and F.P. Noonan. 1983. Mechanism of immune suppression by ultraviolet irradiation in Vivo. I. Evidence for the existence of a unique photoreceptor in skin and its role in photoimmunology. J. Exp. Med. 157:84-98.
Fisher, M.S. 1978. A systemic effect of ultraviolet irradiation and its relationship to tumor immunity. Natl. Cancer Inst. Monogr. 50: 185-88.
Fisher, M.S. 1977. Immunologic Aspects of UV Carcinogenesis, Ph.D. Thesis, University of Utah, School of Medicine, Salt Lake City, UT, 124 pp.
Fisher, M.S., and M.L. Kripke. 1977. Systemic alteration induced in mice by ultraviolet light irradiation and its relationship to ultraviolet carcinogenesis. Proc. Natl. Acad. Sci. USA 74: 1688-92.
Fisher, M.S., and M.L. Kripke. 1978. Further studies on the tumor-specific suppressor cells induced by ultraviolet radiation. J. Immunol. 121: 1139-44.
Fisher, M.S., and M.L. Kripke. 1982. Suppressor T lymphocytes control the development of primary skin cancers in ultraviolet-irradiated mice. Science 216: 133-35.
Greene, M.I., M.S., M.L. Kripke, and B. Benacerraf. 1979. Impairment of antigen-presenting cell function by ultraviolet radiation. Proc. Natl. Acad. Sci. 76:6592-95.
Gurish, M.F., L.K. Roberts, G.G. Krueger, and R.A. Daynes. 1981. The effect of various sunscreen agents on skin damage and the induction of tumor susceptibility in mice subjected to ultraviolet irradiation. J. Invest. Dermatol. 76: 246-251.
Hodges, N.D.M., S.H. Moss, and D.J.G. Davies. 1977. The sensitizing effect of a sunscreen agent, P-aminobenzoic acid, on near UV induced damage in a repair deficient strain of Escherichia coli. Photochem. Photobiol. 26:493-98.
Kligman, L.H., F.J. Akin, and A.M. Kligman. 1980. Sunscreens prevent ultraviolet photocarcinogenesis. J. Am. Acad. Dermatol. 3(a):30-35.
Kligman, L.H., F.J. Akin, and A.M. Kligman. 1982. Prevention of ultraviolet damage to the dermis of hairless mice by sunscreens. J. Invest. Dermatol. 78:181-89.
Kligman, L.H., F.J. Akin, and A.M. Kligman. 1983. Sunscreens promote repair of ultraviolet radiation-induced dermal damage. J. Invest. Dermatol 81:98-102.
Morison, W.L. 1984. The effect of a sunscreen containing para-aminobenzoic acid on the systemic immunologic alterations induced in mice by exposure to UV-B radiation. J. Invest. Dermatol 83:404-8
Noonan, F.P., E.C. De Fabo, and M.L. Kripke. 1981. Suppression of contact hypersensitivity by ultraviolet radiation; an experimental model. Springer Sem. Immunopatho. 4:293-97.
Noonan, F.P., E.C. De Fabo, and M.L. Kripke. 1981. Suppression of contact hypersensitivity by UV radiation and its relationship to UV-induced suppression of tumor immunity. Photochem. Photobiol. 34:683-89.
Noonan, F.P., M.L. Kripke, G.M. Pedersen, and M.I. Green. 1981. Suppression of contact hypersensitivity in mice by ultraviolet irradiation is associated with defective antigen presentation. Immunology 43:527-33.
Pathak, M.A., T.B. Fitzpatrick, and E. Frenk. 1969. Evaluation of Topical agents that prevent sunburn-superiority of para-aminobenzoic acid and its esterin ethyl alcohol. N. Engl. J. Med. 280(26):1459-63.
Snyder, D.S., and M. May. 1975. Ability of PABA to protect mammalian skin from ultraviolet light-induced skin tumors and actinic damage. J. Invest. Dermatol. 65:543-49.
Spellman, C.W., and R.A. Daynes. 1977. Modification of immunological potential by ultraviolet radiation. II. Generation of suppressor cells in short-term UV-irradiated mice. Transplantation 24:120-26.
Stern, R.S., M.C. Weinstein, and S.G. Baker. 1986. Risk reduction for nonmelanoma skin cancer with childhood sunscreen use. Arch. Dermatol. 122:537-45.
Vogel, H.G., H.G. Alpermann, and E. Futterer. 1981. Prevention of changes after UV-irradiation by sunscreen products in skin of hairless mice. Arch. Dermatol. Res. 270:421-28.
Willis, I., and A.M. Kligman. 1970. Amino-benzoic acid and its esters; the quest for more effective sunscreens. Arch. Dermatol. 102:405-17.
Willis, I., J.M. Menter, and H.J. Whyte. 1981. The rapid induction of cancers in the hairless mouse utilizing the principle of photoaugmentation. J. Invest. Dermatol. 76:404-8.
Wulf, H.C., T. Poulsen, H. Brodthagen, and I. Hou-Jensen. 1982. Sunscreens for delay of ultraviolet induction of skin tumors. J. Am. Acad. Dermatol. 7:194-202.
{kind=link}
{kind=link}
{kind=link}
{kind=link}