 Reproduced, with permission, from:
van der Leun, J. C., Y. Takizawa, and J. D. Longstreth. 1989. Human
health. Chapter 2 in the Environmental effects panel report, 11-24.
United Nations Environment Programme (UNEP) report. Nairobi,
Kenya: United Nations Environment Programme.
Reproduced, with permission, from:
van der Leun, J. C., Y. Takizawa, and J. D. Longstreth. 1989. Human
health. Chapter 2 in the Environmental effects panel report, 11-24.
United Nations Environment Programme (UNEP) report. Nairobi,
Kenya: United Nations Environment Programme.
SUMMARY
The increases in ultraviolet-B (UV-B) radiation that will result from depletion of stratospheric ozone are likely to result in a variety of impacts on human health. These range from an increased incidence of cataracts and skin cancer to possible increases in the incidence or severity of certain infectious diseases.
Exposure to ultraviolet radiation has been associated with damage to the cornea, lens and retina of the eye. The principal corneal damage linked to UV exposures is photokeratitis, which appears to be related to acute UV-B exposures. The principal lenticular damage are cataracts. The relationship between UV-B exposure and two forms of cataracts--cortical and posterior subcapsular--appears to be related to cumulative exposure. Both life-time cumulative exposure and annual average exposure are directly related to risk. It has been estimated that a 1% decrease in stratospheric ozone will be accompanied by a 0.6 to 0.8% increase in cataracts. Retinal damage is rare but can occur particularly in those individuals whose lens have been removed in cataract operations.
Ultraviolet radiation is known to affect the immunological defenses of the skin--the first barrier of the body to foreign agents. In tumor systems and with defined antigens, it is clear that UV radiation compromises the ability of the host to immunologically respond either locally (after low doses) or systemically (after relatively higher doses). Preliminary experiments of infectious diseases using animal models have indicated that UV-B can also adversely affect the ability of animals to respond to or contain various infectious agents. Although there are as yet no epidemiologic data to suggest that such effects occur in human populations, nevertheless, animal data suggest that an increase in the severity of certain infections may occur as UV-B fluxes increase due to ozone depletion. In areas of the world where such infections already pose a significant challenge to the public health care delivery systems, the added insult may be significant.
The relationship between UV-B exposure and two forms of non-melanoma skin cancer--basal cell and squamous cell carcinomas--appears to be one of increased risk with increased total lifetime dose to the target cell. Phenotypic characteristics such as skin color can modify the amount of ambient UV that gets transmitted to the target cell, so that fair-skinned individuals are more susceptible than dark-skinned individuals receiving the same ambient exposure. The relationship between UV-B exposure and melanoma skin cancer is probably more complex; it is certainly less well understood. Some data suggest that intermittent severe exposures, i.e., sunburns, are important. Other studies suggest that early exposures (before the ages of 10-14) are of more concern than those acquired later in life. Using dose-response relationships derived from animal experiments and human epidemiologic studies, it is estimated that a 1% decrease in stratospheric ozone will result in a 3% increase in non-melanoma skin cancer, and a lower but still significant increase in melanoma.
INTRODUCTION AND BACKGROUND
UV-B radiation has many influences on the human body. Some are positive, such as the formation of vitamin D3 in the skin, which helps in the maintenance of bone tissue. In phototherapy, doctors utilize the beneficial effects of UV-B radiation on certain skin diseases. On the other hand, many effects of UV-B radiation are damaging, such as sunburn, snowblindness (a form of photokeratitis), cataracts, aging of the skin, and skin cancer. There also are effects which by themselves cannot be called beneficial or damaging, but which influence other reactions of the body. This applies, for instance, to the influences of UV-B radiation on the immune system. UV-B radiation suppresses allergic reactions of the skin, as well as the immune defense against tumours initiated in the skin by UV-B radiation. There are indications of a suppression of the resistance of the skin, and indeed, of the whole organism, against certain infections.
This Chapter will discuss the consequences for human health of an increase of the UV-B radiation in sunlight. The statement that all effects of UV-B radiation would be enhanced in proportion to the increasing irradiance would be too simple. One reason is that for most influences of UV-B radiation the relationship between dose and effect shows non-linear behaviour in certain areas of the dose-response curve. For instance, the formation of vitamin D3 in the skin by UV-B radiation is self-limiting [Holick et al., 1982]. In other cases, adaptation of the skin to UV radiation will modify the effects. Exposure of the skin to UV-B radiation leads to thickening of the most superficial skin layers and to increased pigmentation. This increases the absorption of UV-B radiation in the superficial layers and provides the skin with an efficient protection against sunburn. The hyperproliferation of skin cells involved may, however, also render the skin more susceptible to skin cancer. In some instances, the modifying influences may be so strong that greater doses of radiation lead to smaller effects. Patients with photodermatoses have abnormal reactions of the skin to sunlight, and in many cases to the UV-B radiation in sunlight; the skin will react with papules, itching, swelling, etc. These diseases generally occur less and in less severe forms in the sunny areas of the world than in areas with limited sunlight and dark winters.
Due to such complications and the limited existing knowledge regarding some of the effects of UV-B radiation on man, the overall consequences of an increase of UV-B irradiance on health cannot be predicted in a straightforward matter. For some effects, sufficient data are available to allow quantitative predictions. In other respects, the consequences are rather uncertain. In most cases, something can be learned from comparisons of people living with different exposures to UV-B radiation regardless of any ozone depletion.
The State of the Science section that follows presents both qualitative and quantitative answers to the question of what may be the consequences for human health of stratospheric ozone depletion and its accompanying increase in UV-B. Quantitative estimates of how increased UV-B will affect health are presented for cataracts and non-melanoma and melanoma skin cancers. The issue of the impact on the immune system has been treated qualitatively. It should be stressed, however, that just because there are quantitative estimates for some effects and not for others that this does not imply a ranking of worth. Indeed, the quantitative estimates should be treated very cautiously because they require a number of assumptions. In addition, the effects that can be assessed only qualitatively may well have the potential for a much greater impact.
STATE OF THE SCIENCE
Ocular Damage: Effects on the Cornea, Lens and Retina, and Intraocular Melanoma
Current scientific evidence indicates that exposure to ultraviolet radiation may contribute to the development of a variety of eye disorders [USEPA, 1987; Pitts 1989; Engel et al., 1988; Taylor et al., 1988]. The UV-B portion of the spectrum can damage the cornea, lens and, to a lesser extent, the retina [Pitts, 1989]. Endpoints of concern include cataract, photokeratitis, retinal degeneration, arcus senilis and pterygia.
Damage to the Cornea
The initial response of the eye to exposure to UV-B radiation (280-315 nm) is the condition termed photokeratitis in which the front of the eye, the eyelids and the skin surrounding the eyes become reddened. This condition is commonly seen in skiers and also is called snowblindness. Unlike the skin, the eye does not develop a tolerance to UV but becomes more sensitive with repeated exposures. At wavelengths below 290 nm, damage is principally to corneal epithelium. Between 290 and 315 nm, the corneal stroma and endothelium begin to show damage as well. Doses as low as 0.21 J c[-2] of 308 nm radiation (from a laser) could initiate damage to the cornea; 0.6 J c[-2] at 300 nm produced demonstrable damage in the stroma and endothelium which was not fully repaired eight days post-exposure. Other data indicated that damage from UV radiation at these wavelengths is cumulative and additive among wavelengths [Pitts, 1989].
Damage to the Lens
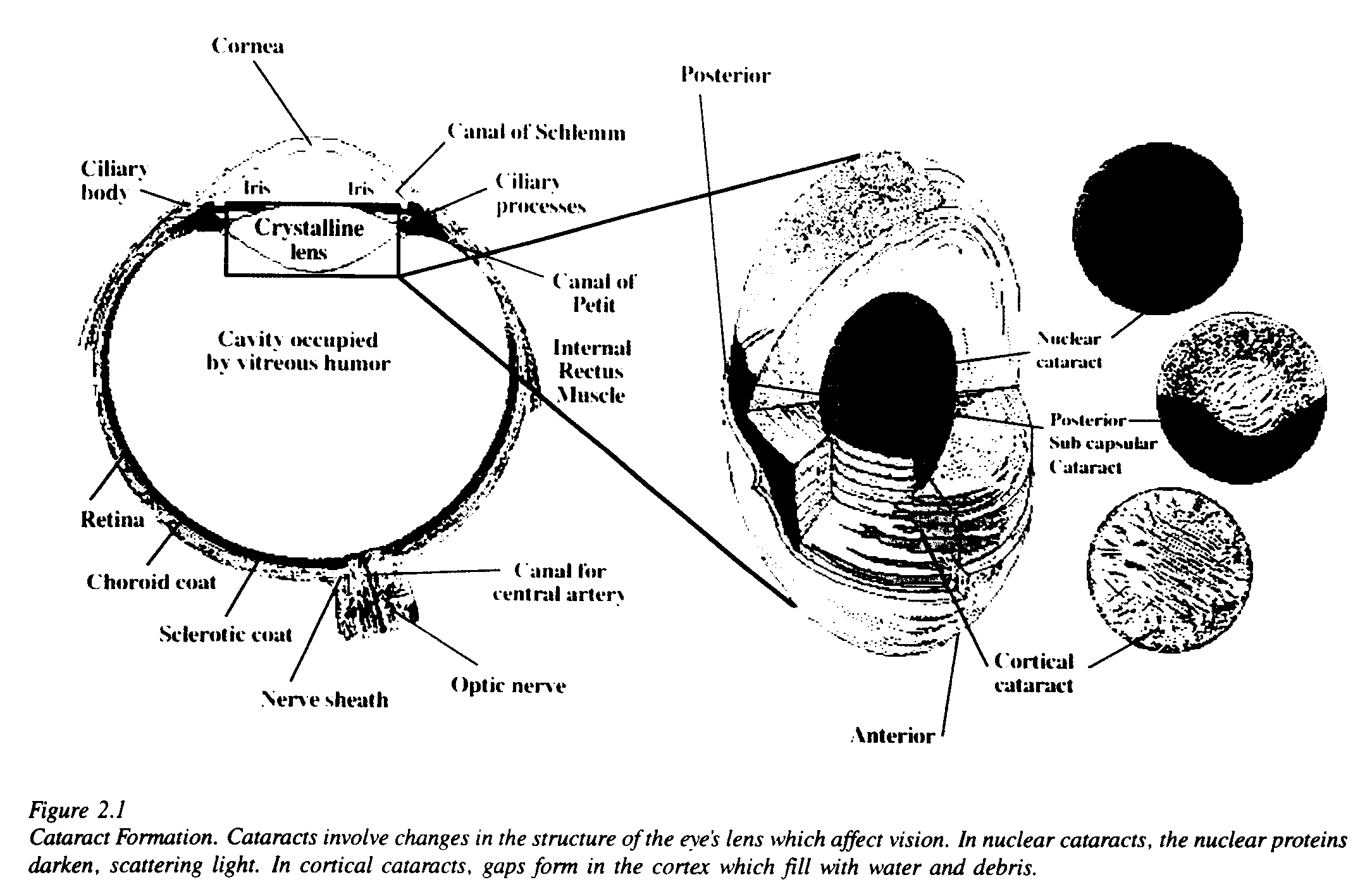
Cataract, a disease of the lens, is the most prevalent form of ocular damage associated with UV exposure. Cataracts are opacities in the lens of the eye which impair vision. There are three major forms of cataracts: nuclear, posterior subcapsular and cortical. In nuclear cataracts, one sees a yellowing of nuclear proteins. In posterior subcapsular cataracts, abnormal and degenerate cells migrate and accumulate at the posterior surface of the lens. In cortical cataracts, gaps form in the cortex and fill with water and debris. Figure 2.1 shows a composite drawing of the eye which indicates the position of the lens and shows how the lens cortex, posterior capsule or nucleus may be affected by a cataract.
In developed countries, corrective surgery can prevent most cataracts from causing blindness; more than 1 million cataract operations were performed in the United States in 1988. Nevertheless, in the United States, cataract remains the third leading cause of legal blindness. In developing countries where such operations are not always available, cataracts result in a much higher incidence of blindness. The World Health Organization (WHO) estimated in 1985 that the greatest single cause of avoidable blindness was cataract. Cataracts were responsible for 17 million cases of blindness, accounting for more than one-half of all the blindness in the world [Maitchouk, 1985].
In both the developed and the developing countries, cataracts represent a significant problem, which is anticipated to grow worse as life expectancy increases. In 1980 almost one-half of the world's 260 million elderly resided in the developed nations, whereas by the year 2000, an estimated two-thirds of the elderly (400 million) will reside in the now developing nations [Maitchouk, 1985]. Thus, cataract is expected to grow as a public health problem in the developing countries.
The exact mechanism of the formation of a cataract is still unknown, although it is clearly multifactorial. Epidemiological studies [USEPA, 1987; Taylor et al., 1988, Bochow et al., 1989] suggest that some cataracts, particularly cortical and posterior subcapsular cataracts [Taylor et al., 1988; Bochow et al., 1989], are etiologically related to exposure to solar radiation, specifically UV-B [Hollows and Moran, 1981]. Studies in animals suggest that the active portion of the solar spectrum lies in the UV-B region [Pitts et al., 1986]. The cataracts induced in these studies were anterior subcapsular [Pitts et al., 1977]. Ultraviolet-A radiation and other causes, such as nutritional deficiency, also may contribute to cataract formation.
Table 2.1 presents the relationship between cataract incidence and amount of exposure to sunlight in Nepal [Brilliant et al., 1983]. Even more persuasive, however, is the finding in a recent epidemiologic study of occupationally exposed individuals that the incidence of cortical cataracts appears to be directly related to exposure to UV-B. A doubling of cumulative exposure increased the risk of cortical cataract by a factor of 1.6. Nuclear cataracts showed no such increased risk with UV-B exposures. When individuals were ranked as to their average annual UV-B exposure, those in the upper quarter of the ranking had a 3.3 fold increased risk of cortical cataract compared to those in the lowest quarter [Taylor et al., 1988]. A second recent case-control study examined the relationship between posterior subcapsular cataract and UV-B and found that cases had a 23% higher cumulative exposure to UV-B [Bochow et al., 1989]
Given this evidence, reduction in the ozone layer and its associated increase in the amount of UV-B reaching the earth's surface will likely increase the incidence of cataracts. On the basis of earlier epidemiologic data [Hiller et al., 1983], the U.S. Environmental Protection Agency estimated that for every 1% decrease in stratospheric ozone, there will be between a 0.3 and 0.6% increase in cataracts. More recently, the work of Taylor and his colleagues [1988] has indicated that for every 1% decrease in stratospheric ozone there will be a 1.2% increase in cortical cataracts. Given that nuclear cataracts account for about 30% of all cataracts and that posterior subcapsular cataracts account for about 40% and apparently have approximately the same dose-response relationship as that observed for cortical cataracts, the EPA estimates might need to be revised upward slightly to 0.6-0.8% [Taylor, 1989]. Application of this range of estimated increases (0.6-0.8%) in cataracts to the number of cases of cataract blindness estimated for 1985 [17 million; Maitchouk, 1985] suggests that if the same population had lived with the ozone layer reduced by 1%, there would have been between 100,000 and 150,000 additional cases of cataract-induced blindness.
Damage to the Retina
UV reaching the retina causes both functional and morphological damage. Lenses that have been removed (aphakic), such as after a cataract operation, provide clear evidence of this occurence. The threshold for damage to the rabbit eye at 300 nm was 0.23 J [-2], A similar value (0.36 J [-2]) was found for the primate eye at 325 nm (not UV-B); the aphakic threshold was about ten times lower. More research is needed to determine how these findings might apply to retinal damage in exposed human populations.
Intraocular Melanoma
Sunlight exposure has recently been implicated as a risk factor for intraocular melanoma in a study that determined a latitude gradient for this tumour. Individuals born in the southern United States showed nearly a three-fold increase in risk. Blue-eyed individuals comprised the phenotype with the largest risk. Complexion and hair colour were not important risk factors [Tucker et al., 1985]
Immunologic Effects/Infectious Diseases
Ultraviolet radiation has profound effects on the immune system, particularly that of the skin. At ambient environmental levels, animal models have shown that UV radiation reduces the ability of the cell-mediated arm of the immune system[1] to respond adequately to foreign substances, known as antigens [DeFabo and Kripke, 1979]. The skin often is the first point of contact with many foreign substances including infectious agents. In these instances, the skin's immune response is the body's first line of defense. However, when foreign substances (antigens) are given via the skin to UV-treated animals, they induce tolerance rather than inducing a protective response (immunity). The reason for this failure appears to be due to an increase in or elicitation of the activity of a class of lymphocytes termed suppressor T-cells (Ts cells). This occurs possibly in conjunction with the failure of a second part of the immune system: the antigen presenting cell [DeFabo and Noonan, 1983; Granstein, 1984; Baadsgaard, 1987]. Normally, if an antigen is introduced via the skin, a cellular immune response to it would be generated via the interaction of antigen-presenting cells and a class of lymphocytes termed effector T-cells. In UV-exposed skin, the loss of one type of antigen-presenting cell type--the Langerhans cell--is associated with the subsequent appearance of Ts cells [Elmet et al., 1983]. Such cells normally serve a regulatory function, preventing an animal from making inappropriate immune responses such as responding to its own proteins.
In the case of UV-treated skin, the Ts cells prevent the animal from responding to antigens given at the same time or shortly after these cells appear in the skin. It is for this reason that in animal models, UV radiation-induced tumours are not controlled by the skin's immune system; they are not recognized as foreign. This observation has been explored in some detail in the context of the induction and development of tumours by UV radiation [Fisher and Kripke, 1974, 1981; Kripke, 1984]. However, it has important implications not only for carcinogenic processes but also for many infectious disease systems, where the inability of the host to respond to infectious agents in the skin may allow such infections to become established.
The skin cancer effects of UV radiation occur mainly in the white-skinned races. Most information suggests this is because the greater pigment (melanin) content of dark skin blocks UV radiation, preventing it from harming the cells of the skin. Were this same reasoning applied to the immune system effects, the conclusion would be that only the light-skinned populations would be at risk for adverse effects on the immune system. There are a limited number of studies that address this issue. However, what little has been reported indicates that the effects of UV radiation on the immune system occurs irrespective of skin colour. Thus, in humans, the effect of UV radiation on the antigen presenting cell has been observed at very low doses not only in Caucasians but also in Asians and Aborigines [Scheibner, 1987]. On the basis of this information, the impacts on the immune response and its role in the control of infectious diseases may not be restricted to the white populations.
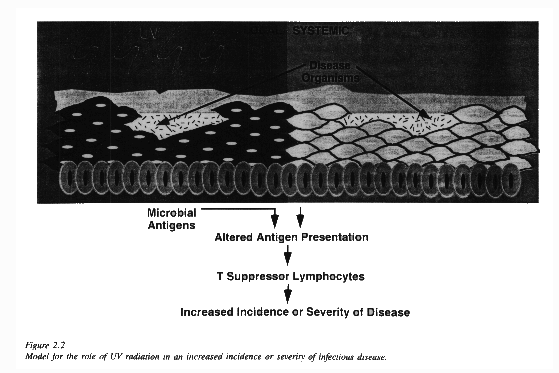
There are a large number of diseases that might be affected by UV-induced immunosuppression. At a minimum, these include all those diseases which have a stage involving the skin, and to which cell-mediated (as opposed to antibody-mediated) immunity is important. Examples include: 1) measles and other viral diseases that elicit a rash such as chicken pox and herpes, 2) parasitic diseases introduced via the skin such as malaria and leishmaniasis, 3) bacterial diseases such as tuberculosis (T-cell immunity is important), and 4) fungal infections such as candidiasis. Studies of animal models of some of these diseases confirm the potential for an effect of UV-B on the development of infectious disease processes [Howie, 1986a,b; Giannini, 1987; Taylor, 1988]. Figure 2.2 presents a model of how ultraviolet radiation is thought to interact with skin inducing Ts cells resulting in specific non-responsiveness to infectious agents that subsequently gain entry into the skin.
Figure 2.2 deals only with impaired local immunity (i.e., at the site of irradiation), which occurs following doses of UV that could be received in a normal sun-exposure scenario such as a week-long sunny vacation. Larger doses (i.e., several months of daily sunny exposure) could possibly induce impaired systemic immunity so that introduction of any organism might be met by a diminished host response.
Brief descriptions follow of several diseases (herpes leishmaniasis, malaria, and bacterial and fungal infections) where preliminary results have shown that UV radiation has an impact. With the exception of herpes virus infections where UV-B irradiation has been shown to elicit reoccurrence of the virus in man, most of the data come from studies in animals. There are other diseases which also may be candidates for an impact of increased UV radiation by virtue of one of the traits identified above. Brief descriptions of two of these--tuberculosis and leprosy--are given below. However, experimental evidence to examine these hypothese needs to be gathered.
Herpes Viruses
Human herpes viruses comprise a class of viruses that include the varicella-zoster virus (the cause of chicken pox and herpes zoster), the herpes simplex virus (cold sores and genital herpes), cytomegalovirus, and the Epstein-Barr virus (Burkitt's lymphoma and infectious mononucleosis). These viruses have as a common characteristic the ability to exist in a carrier state which can last for extended periods of time. In the carrier stage, the infection has no symptoms, but a variety of stimuli can cause reactivation. For herpes simplex viruses, reoccurrences have been noted after fever, trauma, emotional upsets, gastrointestinal disorders, and exposure to UV. In humans, both multiple sub-erythemal doses (10 exposures, equivalent to 5 or 6 MED[2]) or single doses equivalent to a mild sunburn (3 MED) have been shown to reactivate latent herpes simplex virus infections [Spruance, 1985; Perna et al., 1988]. The mechanism leading to reactivation of herpes in humans is unknown. Studies in mice, however, indicate that UV irradiation of mice at the site of cutaneous infection with herpes led to the development of Ts cells that decreased the activity of those T cells that would normally react to herpes-infected cells [Aurelian et al., 1988].
Leishmaniasis
Leishmaniasis is a parasitic protozoan infection transmitted by the sand-fly. Human leishmaniasis is caused by several different species and sub-species of protozoa and is present on every continent except Australia and Antarctica. The World Health Organization has estimated that there are 400,000 new cases of leishmaniasis each year [UNDP, 1980], with the majority of cases being found in Africa and Latin America. WHO also concluded that this was a gross underestimate.
In studies using a broad spectrum UV lamp, a dose of 150 J [-2] UV-B reduced the size of the lesion-induced leishmania but did not change the distribution of leishmania parasites in an animal model of the skin [Giannini. 1986]; 79% of the lamp's output was in the 280-320 nm range, and the remainder was above 320 nm. In a subsequent study, Giannini and DeFabo [1987] used monochromatic UV-B light at 290 nm or 320 nm. They found that 720 J [-2] of 290 nm (given in five, 1-hour exposures of about 145 J [-2]) appeared to promote the dissemination of parasites but did not modify lesion development. In contrast, five exposures to 330 J [-2] of 320 nm UV-B (total dose = 1650 J [-2]) reduced lesion size but did not appear to promote dissemination. These observations lead to the hypothesis that exposure to sunlight, particularly as it becomes enriched with the shorter wavelengths of UV-B due to ozone depletion could have an adverse impact on skin diseases such as leishmaniasis. These observations suggest that exposure to sunlight could result in a suppressed immune response to the parasite and predispose the individual to a more severe disease.
Malaria
Malaria in humans is caused by one of four Plasmodium parasites and is transmitted by the mosquito. The immune responses against the four strain-specific malarial infections differ. Generally, malarial infection induces both humoural and cell-mediated immunity [Braunwald, 1987]. Numerous studies have suggested that suppressor T-cells are normally induced during malarial infection, but their presence has not been demonstrated conclusively. Preliminary studies in a mouse model of malaria have indicated that UV irradiation at low doses can impair the ability of the host to control the infection [Taylor, pers. comm.]. However, more work in both human and animal systems is needed to determine if UV radiation may have a role in the development of or response to malaria in humans.
Bacterial and Fungal Infections
Bacterial and fungal cells are in constant residence on the skin and are the cause of the majority of skin infections. As a general rule, such infections are relatively minor, but in immunosuppressed individuals they can be a major cause of morbidity. Some of the more commonly found microbial inhabitants of the skin are the yeast Candida albicans and the bacteria Staphlococcus aureus and Escherichia coli. Preliminary data have shown that animals given high doses of ultraviolet radiation (15 KJ [-2]) have difficulty in clearing either S. aureus, E. coli, or C. albicans when given an intravenous dose [Chung et al., 1988].
Tuberculosis and Leprosy
Tuberculosis is increasingly becoming a problem in the United States and many other areas of the world. In the United States in 1987, there were 22,517 cases of tuberculosis reported to the Centers for Disease Control, an increase of about 3% compared to 1986 [CDC, 1988]. After chicken pox and food poisoning (salmonellosis), tuberculosis had the highest number of cases reported of any infectious disease not classified as sexually transmitted. Immunity to the mycobacteria that causes tuberculosis is T-cell dependent, has a cutaneous phase, and is best elicited via immunization in the skin. Therefore, tuberculosis has many of the same qualities that identify diseases whose immunity may be compromised by UV radiation. Preliminary studies in an animal model of immunity to tuberculosis (immunization with Bacillus Calmette-Guerin) also suggest that moderate doses of UV radiation may impair development of an immune response and retard recovery [Kripke, 1989].
Leprosy also is caused by a mycobacterium and has many other characteristics which are similar to those of tuberculosis. T-cell mediated immunity is important in a host's response to leprosy and there is a cutaneous stage of the disease. It seems plausible to consider this too as a candidate disease which may be affected by increases in UV radiation as a result of ozone depletion.
Other Concerns: Vaccination Programs
The potential effects of increased UV-B on infectious diseases include not only direct increases in morbidity and mortality but via interference with vaccination programs as well. If an individual is immunized through UV-treated skin, the potential exists for that treatment to render the individual more susceptible rather than less susceptible to the administered antigen. This could have serious ramifications for vaccination programs in lesser developed countries where the control of certain infections (e.g., measles) depends on effective vaccination.
Skin Cancer
Skin cancer is the most common form of cancer in white populations. There are two types of skin cancer: 1) non-melanoma skin cancer (NMSC) which affects the keratinocytes in the skin, and 2) cutaneous malignant melanoma (CMM) which affects the pigment-producing cells of the skin. The risk of developing or dying from these cancers varies significantly by race. American blacks have an incidence rate of nonmelanoma skin cancer that is 1% of the incidence rate for whites [Scotto, 1989], and an incidence rate of CMM which is 10% of that of whites [NCI, 1988]. During the period 1977 to 1985, CMM increased 57% in whites and decreased 10% in American blacks. Their average age-adjusted incidence rates (adjusted to the 1970 U.S. standard population) per 100,000 were 8.3 and 0.8 respectively [NCI, 1988]. Similar data for Asians in the United States are not readily available. However, for the period from 1971 to 1977, the age-adjusted melanoma mortality in Japan was about 10% of that observed for the 1973 to 1977 period in the United States (0.16 versus 1.9 per 100,000; age adjusted to the 1970 Japanese population census) [Takizawa, 1987; NCI, 1988].
Non-Melanoma Skin Cancer[3]
There are two main types of non-melanoma skin cancers: basal cell carcinoma (BCC) and squamous cell carcinoma (SCC). The data currently gathered on these tumours are highly variable. In many instances, these tumours are removed in physicians' offices, and histopathology to identify type and/or reporting to a cancer registry is not performed. In registries that record these tumours, a few include only SCC, whereas most record a combination of BCC and SCC incidence under the designation of non-melanoma skin cancer. Where good medical care is available, the overall mortality is less than 1%. Although BCC typically represents 80% of non-melanoma skin cancer, the mortality is mainly due to SCC.
Squamous cell carcinomas have a convincing and clear-cut relationship to UV-B radiation. The causative role of sunlight is supported by the following observations: 1) SCC occurs predominantly on the most sun-exposed parts of the skin, face, neck and hands; and 2) in comparable populations, the incidence of SCC is highest in locations with the most sunlight. SCC also occurs predominantly in fair-skinned people, presumably due to a lack of protective pigment. The risk of developing SCC is strongly related to the cumulative dose of sunlight received throughout life. Several experimental animals develop SCC from exposure to UV radiation. A wealth of data is available, particularly for mice. De Gruijl et al. [1983] determined the dose-effect relationship for SCC induced by UV-B radiation. The carcinogenic effect could be described in mathematical terms as a power function of the doses of UV-B radiation regularly received by the animals. The basic same dose-effect relationship can be traced back in human epidemiological data [Fears et al., 1977]. This adds confidence to the assumption underlying the use of experimental data in this connection; the processes of photocarcinogenesis in mouse and human are generally the same.
The wavelengths responsible for UV-carcinogenesis in mice are predominantly in the UV-B range [Roffo, 1934; Blum, 1959], indicating that a depletion of stratospheric ozone has the potential to increase the incidence of skin cancer. As discussed in Chapter 1, one piece of information required for a quantitative consideration of this effect is an action spectrum for UV-carcinogenesis. The action spectrum gives the carcinogenic effectiveness of UV radiation as a function of wavelength. Until recently, such a relationship has not been available, and investigators had to work with hypothetical action spectra. It was assumed, for instance, that the unknown action spectrum would be the same as that for UV-erythema, the reddening of the skin by UV radiation; the latter action spectrum has a pronounced peak in the UV-B range. Support for this assumption came from the work of Cole et al. [1986]. These investigators found that experimental data on photocarcinogenesis in mice could be described satisfactorily if it were assumed that the action spectrum for UV-carcinogenesis would be the same as that for UV-induced edema in mouse skin. The latter action spectrum was very similar to that for UV-erythema in human skin.
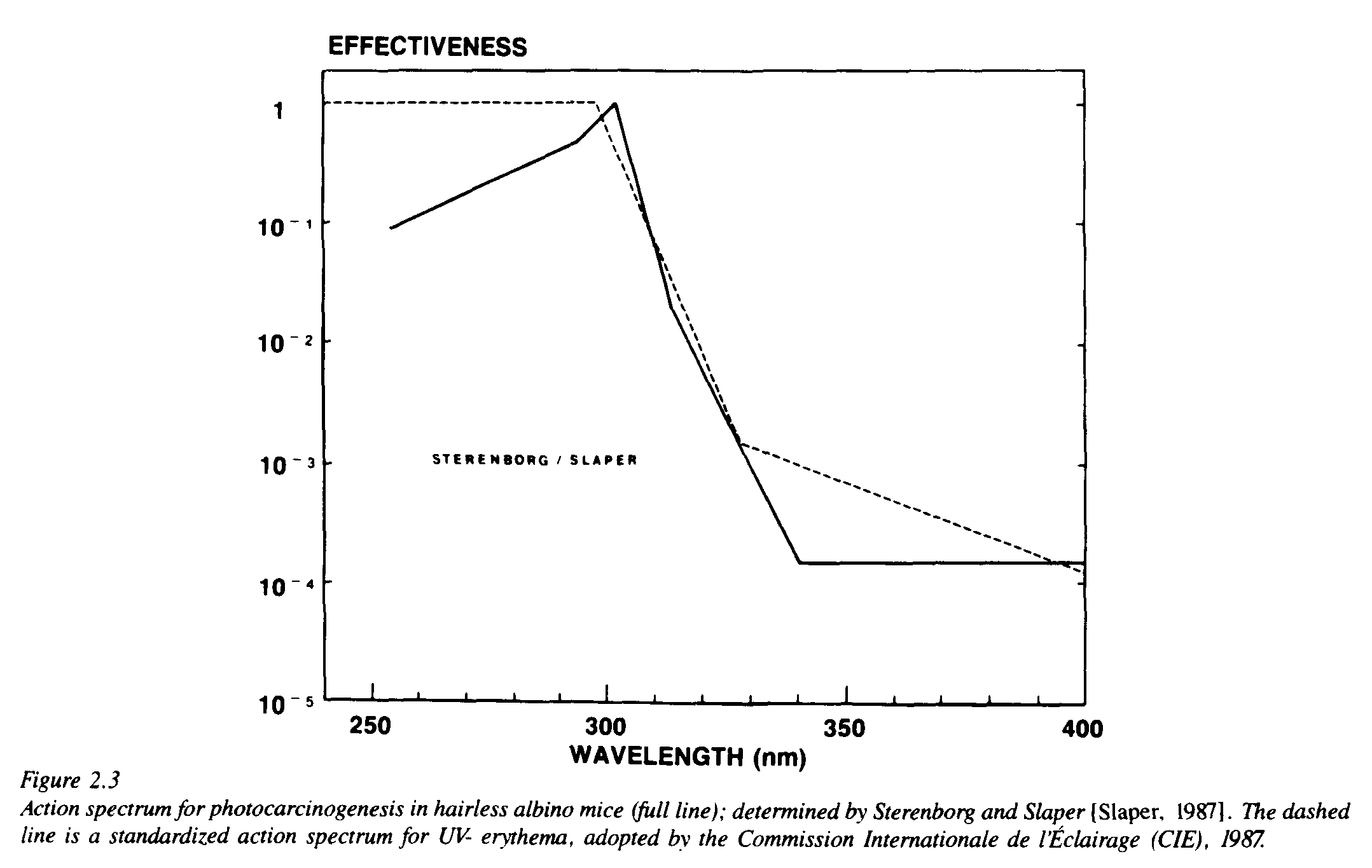
A programme to determine the action spectrum for UV-carcinogenesis in mice was carried out by Sterenborg and van der Leun [1987] and Slaper [1987]. These investigators exposed groups of hairless mice to UV radiation from eight different combinations of lamps and filters; from the experimental results they computed an action spectrum for UV-carcinogenesis (Figure 2.3). The action spectrum consists of a limited number of straight line segments, which indicates that it still is a crude approximation. The curve is best defined in the UV-B range, where a comparatively greater amount of experimental information is available. In this wavelength range, which is important from the viewpoint of ozone depletion, the action spectrum shows a close correspondence to that for UV-erythema. This tends to support, at least in general, earlier predictions based on less complete data.
With both the dose-effect relationship and the action spectrum determined, more knowledge is available on the induction of squamous cell carcinoma by UV radiation than on any other potential consequence of ozone depletion. This has led to comparatively well-developed estimates of the increase of SCC with a decrease in stratospheric ozone. The estimates, which are being checked and refined, may also be of help in developing projections for other biological consequences. One conclusion is that the number of patients with SCC will increase more sharply than the rate at which the amount of ozone decreases. This is expressed in an "amplification factor," giving the percentage increase of the incidence caused by a 1% decrease of ozone. This percentage exceeds 1 as a result of the combined action of two amplifications: radiation amplification and biological amplification.
The radiation amplification, discussed in Chapter 1, represents the fact that a 1% decrease of ozone gives a stronger increase of the effective irradiance at the earth's surface. It depends on the wavelengths involved in the biological effect under consideration, because the increased penetration in the case of ozone depletion is wavelength dependent; it is greatest for the shortest wavelengths in the solar spectrum.
Figure 2.3 shows that in this wavelength range, especially in the wavelengths just above 300 nm, the carcinogenic effectiveness depends steeply on wavelength. This implies that an increase of irradiance in these wavelengths will have a comparatively large effect. Recent computations show that, with the Sterenborg-Slaper action spectrum, the radiation amplification factor has a value of 1.6 [Kelfkens and de Gruijl, 1989]. This is close to the values calculated before, on the basis of hypothetical action spectra; the values usually ranged between 1.7 and 2.0.
The biological amplification represents the fact that a 1% increase of the effective irradiance leads to a greater increase of the incidence of SCC. This amplification does not relate to the wavelengths responsible for carcinogenesis, but is due entirely to the steepness of the power relationship between the doses of UV radiation regularly received and the incidence of skin cancer. It may be concluded from epidemiological data in the United States [Scotto et al., 1981] that the biological amplification factor for SCC has a value of 2.9 [Slaper et al., 1986].
If the two amplifications are taken together, these data indicate that a 1% decrease of ozone will give a 1.6% increase of the effective irradiance; this will lead to an increase of the incidence of SCC by 1.6 x 2.9 = 4.6%. As SCC is a long-term reaction of the skin to chronic irradiation, it will take several decades for this increase to be fully realized.
For basal cell carcinoma, the situation is less clear. Although the body site distribution of BCC is similar to that for SCC, BCC has a latitudinal gradient that is less extreme. Furthermore, mice under UV lamps rarely develop BCC, so that an action spectrum specific to this tumour has not been determined. The clinical and epidemiological analogies between SCC and BCC are usually taken as an indication that BCC is also a consequence of solar UV-B radiation. If an additional assumption is made that the induction of BCC has the same action spectrum as the induction of SCC, it is possible to calculate the amplification factors. On this basis, the biological amplification factor for BCC becomes 1.7, and the overall amplification factor 1.6 x 1.7 = 2.7. It must be stressed that this latter value is more uncertain than the amplification factor for SCC. The amplification factors given here are in good agreement with those derived by others in a different way, but basically from the same data [Scotto et al., 1981].
The greater uncertainty is found in the most frequent form of non-melanoma skin cancer: basal cell carcinoma. The best-founded numbers refer to the more aggressive form: squamous cell carcinoma. For areas where only combined data on non-melanoma skin cancer are available, a value of 3 for the overall amplification factor appears to be the most appropriate. That means a 1% decrease of ozone will ultimately lead to a 3% increase of the incidence of non-melanoma skin cancer.
These calculations of increases of incidence to be expected for a 1% depletion of ozone form the basis for calculating the consequences of larger depletions. The depletion of total column ozone expected has varied over the years, with the improvement of atmospheric models. The expected depletions also will be influenced by real reductions of the use of CFCs, as agreed in the Montreal Protocol or by more stringent future limitations. These are sound reasons for giving the biological changes predicted "per percent of ozone depletion." In this way, the calculations of biological consequences can be readily adjusted to new data and new scenarios.
Should total column ozone be reduced by 5%, the carcinogenically effective UV-B irradiance would increase by 5 x 1.6 = 8%. In the long run, this would lead to an increase of BCC by 14% and of SCC by 25%. The number of new patients affected may be estimated from data available on the present incidences. For the United States where comparatively good data are available, it is estimated that there are 500,000 new patients with non-melanoma skin cancer per year, 80% of these having BCC and 20% SCC. The increases calculated for a 5% depletion of ozone would come down to 56,000 additional patients with BCC and 25,000 with SCC, or a total of 81,000 additional patients with non-melanoma skin cancer per year. Fewer data are available on the incidence of NMSC in many other parts of the world. It appears that a conservative estimate of the total number of non-melanoma skin cancer patients worldwide would be three times that in the United States. This would also lead to a three-fold increase, or about 240,000 additional patients with non-melanoma skin cancer per year worldwide.
The quantitative predictions for the incidence of non-melanoma skin cancer are based on comparatively solid data. However, they also are based on the assumption that the behaviour of human populations will not change. Human behaviour has important influence on the doses of UV radiation received, and a general change of behaviour could have a profound influence on the incidence of skin cancer. The rise of incidence expected from ozone depletion will come superimposed on any change of the incidence resulting from other causes.
Cutaneous Malignant Melanoma
Incidence rates of cutaneous malignant melanoma among white-skinned races throughout the world are rising at an alarming rate. During the period from 1974 to 1986, CMM incidence in the U.S. increased at an average yearly rate of 3 to 4%. For more than a decade. there has been serious concern that CMM is at least partially caused by exposure to sunlight and, in particular, by exposure to ultraviolet-B radiation [NAS, 1979, 1980, 1982]. However, several aspects of the scientific information about CMM have puzzled researchers and have contributed to uncertainty about the relationship of CMM to solar radiation and to UV-B. A brief overview of the biology of CMM is given below. followed by summaries of the information from human studies and animal models that must be considered in evaluating this relationship.
Cutaneous malignant melanoma is the result of the neoplastic transformation of melanocytes which are the pigment-producing cells in mammalian epidermis. Four different types of CMM have been evaluated for their relationships to sun exposure:
The two most common forms are SSM and LMM. It is thought that the role of sunlight in the induction of these tumours is very different. LMM shows a relationship similar to the cumulative dose relationship with BCC and SCC, whereas the relationship between solar exposure and SSM (and the other forms as well) may be related to peak exposures or possibly exposures early in life.
Evidence supporting a relationship between CMM and solar radiation, in particular UV-B, includes the following points [USEPA, 1987]:
In the past, the controversy with regard to the relationship between solar exposure and CMM other than LMM revolved around several observations that apparently contradict the kind of direct relationship between solar exposure and tumour development observed with non-melanoma skin cancer. Some of the key points include the following:
Some of these issues have been resolved. For example, there are now two animal models (discussed below). At least one study [Pearl and Scott, 1984] has reanalyzed the site distribution data and suggests that on a surface area basis, CMM does occur on the more sun exposed areas. Clearly, however, CMM does not show the same propensity to develop on the head and hands that is demonstrated by non-melanoma skin cancer.
The U.S. Environmental Protection Agency completed a review of the available data and literature to assess the relationship of CMM to UV-B exposure [USEPA, 1987]. The salient conclusions of that review were the following: 1) the older literature, taken in conjunction with some very recent case-control epidemiologic studies, clearly indicated a role for sunlight in the etiology of all CMM, not just LMM; 2) based on animal studies, the active portion of the solar spectrum was most likely to be in the UV-B region, implicating UV-B in non-melanoma etiology and immunosuppression and on the sensitivity of XP patients; and 3) the dose-response relationships for incidence and mortality were estimated to be such that for every 1% decrease in stratospheric ozone there would be up to a 2% increase in CMM incidence and between 0.3 and 2% increase in CMM mortality.
Until recently, there were no animal models suitable to investigate the induction of melanomas by UV radiation. Thus, there was no possibility to check experimentally the suggestions based on epidemiology about the involvement of sunlight in the generation of melanomas, and no possibility to determine an action spectrum or a dose-effect relationship. Until such information is available, a high degree of uncertainty remains on any quantitative projections of an influence of ozone depletion on the incidence of melanomas. Recently, however, two promising animal models were found. Woodhead et al. [1988] reported on a small hybrid fish that developed melanomas after exposure to UV-B radiation. Ley [1988] found that UV-B radiation could induce melanomas in the marsupial Monodelphis domestica. Both models are new and publications have not yet appeared in print. It will take time before useful quantitative information is yielded by such models. But the emergence of these models supports the impression that UV-B radiation has a role in the induction of human melanomas.
Linkages
Global Warming
The relationships between weather parameters and human health are very complex, making it difficult to predict exactly what the impacts of global warming will be on health end-points. Potential concerns that have been identified, however, include the following [USEPA, 1989]:
Infectious diseases comprise a large area of potential overlap between the potential effects of global warming and stratospheric ozone depletion on human health. If the immunosuppressive effects of increased UV have an impact on infectious diseases, then this could have an additive or even synergistic effect on vector-borne diseases or perinatal infections. The two impacts could result in an increased incidence driven by climate change, occurring in populations made more susceptible by UV. Vaccination programs designed to deal with these infections could be compromised if vaccines are given to individuals with a high UV exposure. In addition, in the more northern countries where global warming will permit individuals to get out in the sun earlier in the season, the potential exists that a greater UV-B dose will be received under conditions of ozone depletion than would be received if the ozone level were maintained.
Air Pollution
Preliminary data suggest that stratospheric ozone depletion may result in increased levels of tropospheric ozone and other photochemical oxidants. Should this occur, asthmatics and other individuals who are sensitive to ozone levels are likely to encounter air pollution levels injurious to their health. The same prediction has been made vis a vis global warming. Thus, air pollution will be exacerbated by both global warming and ozone depletion thereby suggesting that a major interaction will be an increase in respiratory problems.
Animal Health
UV-B exposure is thought to be responsible for squamous cell cancer of the eye in Herefords and other cattle and has been shown to exacerbate infectious bovine keratoconjunctivitis (IBK) [Kopecky et al., 1979, 1980]. Thus, increases in UV are likely to increase these problems. Most animals, because of their heavy coats. are not likely to suffer the immunosuppressive effects of UV, but it is possible that global warming may change patterns of animal infectious disease via changes in the prevalence and distribution of insect vectors [Stem et al., 1988]. IBK is spread by the face fly; it could experience an augmentative impact both from ozone depletion and from global warming.
REFERENCES
Baadsgaard, O., C.H. Wulf, G.L. Wantzin, and K.D. Cooper, UVB and UVC, but not UVA, potently induce the appearance of T6-DR+ antigen-presenting cells in human epidermis, J. Invest. Dermatol., 89, 113-118, 1987.
Blum, H.F., Carcinogenesis by Ultraviolet Light, Princeton University Press, Princeton, New Jersey, 1959.
Bochow, T.W., S.K. West, A. Azar, B. Munoz, A. Sommes, and H.R. Taylor, Ultraviolet light exposure and the risk of posterior subcapsular cataract, Arch. Ophthalmol., 107, 369-372, 1989.
Braunwald E., K.J. Isselbacher, R.G. Petersdorf, J.D. Wilson, J.B. Martin, and A.S. Fauci (eds.), Harrison's Principles of Internal Medicine, 11th Edition, McGraw-Hill. 1987.
Brilliant, L.B., N.C. Grasset, R.P. Pokhrel, A. Kolstal, J.M. Lepkowski, G.E. Brilliant, W.N. Hawks, and R. Pararajasegaram, Associations among cataract prevalence, sunlight hours and altitude in the Himalayas, Am. J. Epidemiol., 118, 250-264, 1983.
Cano, R.J. and J.S. Colome, Microbiology, West Publishing Co., 1986.
Centers for Disease Control (CDC), Summary of notifiable diseases: United States, 1987, MMWR, 36, 3, 1988.
Chung, H-T, D.G. Lee, S.Y. Lim, and R.A. Daynes, UVR-exposed animals exhibit an enhanced susceptibility to bacterial and fungal infections, Abstract, J. Invest. Dermatol., 90, 552, 1988.
Cole, C.A., P.D. Forbes, and R.E. Davies, An action spectrum for photocarcinogenesis, Photochem. Photobiol., 4, 275-284, 1986.
DeFabo, E.C. and M.L. Kripke, Dose-response characteristics of immunologic unresponsiveness to UV-induced tumors produced by UV irradiation of mice, Photochem. Photobiol., 30, 385-390, 1979.
DeFabo. E.C. and F.P. Noonan, Mechanisms of immune suppression by ultraviolet irradiation in vivo, Part I: Evidence for the existence of a unique photoreceptor in skin and its role in photoimmunology, J. Exp. Med., 157, 84-98, 1983.
De Gruijl, F.R. and J.C. van der Leun, A dose-response model for skin cancer induction by chronic UV exposure of a human population, J. Theor. Biol., 83, 487-504, 1980.
De Gruijl, F.R., J.B. van der Meer, and J.C. van der Leun, Dose-time dependency of tumor formation by chronic UV-exposure, Photochem. Photobiol., 37, 53-62, 1983.
Elmets, C.A., P.R. Bergstresser, R.E. Tigelaar, P.J. Wood, and J.W. Streilein, Analysis of the mechanism of unresponsiveness produced by haptens painted on skin exposed to low dose ultraviolet radiation. J. Exp. Med., 158, 781-794, 1983.
Engel, A., M.L. Johnson, and S.G. Haynes, Health effects of sunlight exposure in the U.S., Arch. Dermatol., 124, 72-79, 1988.
Fears, T.R., J. Scotto, and M.A. Schneiderman, Mathematical models of age and ultraviolet effects on the incidence of skin cancer among whites in the United States, Am. J. Epidemiol., 105, 420-427, 1977.
Fisher, M.S. and M.L. Kripke, Suppressor T lymphocytes control the development of primary skin cancers in ultraviolet-irradiated mice, Science, 216, 1133-1134, 1981.
Fisher, M.S. and M.L. Kripke, Antigenicity of murine skin tumors induced by ultraviolet light, J. Natl. Cancer Inst., 53, 1333-1336, 1974.
Giannini, M.S.H., Suppression of pathogenesis in cutaneous leishmaniasis by UV irradiation, Infect. Immunol., 51, 838-843, 1986.
Giannini, S.H. and E.C. DeFabo, Abrogation of skin lesions in cutaneous leishmaniasis, in Leishmaniasis: The First Centenary (1885-1985) New Strategies for Control, D.T. Hart (ed.), NATO ASI Series A: Life Sciences, Plenum Publishing Company, London, 1987.
Glassroth J.G., A.G. Robins, and D.E. Snider, Tuberculosis in the 1980's, New Eng. J. Med., 302, 14411450, 1980.
Granstein, R., A. Lowy, and A. Green, Epidermal antigen-presenting cells in activation of suppression: Identification of a new functional type of ultraviolet radiation resistant epidermal cell, J. Immunol., 132, 563-565, 1984.
Hiller, R., R.D. Sperduto, and F. Ederer, Epidemiologic associations with cataract in the 1971-1972 National Health and Nutrition Survey, Am. J. Epidemiol., 118, 239-249, 1983.
Holick, M.F., J.A. MacLaughlin, J.A. Parrish, and R.R. Anderson, The photochemistry and photobiology of vitamin D3. pp. 195-218 in The Science of Photomedicine, J.D. Regan and J.A. Parrish (eds.), Plenum, New York. 1982.
Hollows, F.C., and D. Moran, Cataract--the ultraviolet risk factor, Lancet, 2, 1249-1250, 1981.
Howie, S., M. Norval, and J. Maingay, Exposure to low-dose ultraviolet radiation suppressed delayed-type hypersensitivity to herpes simplex virus in mice, J. Invest. Dermatol., 86, 125-128, 1986a.
Howie, S.E.M., M. Norval, J. Maingay, and J.A. Ross, Two phenotypically distinct T-cells (Lyl+2- and Lyl-2+) are involved in the ultraviolet-B light-induced suppression of the efferent DTH response to HSV-1 in vivo, J. Immunol., 58, 653-658, 1986b.
Kelfkens, G. and F.R. de Gruijl, Personal communication, 1989 (to be published).
Kopecky, K.E., G.W. Pugh, D.E. Hughes, G.D. Booth, and N.F. Chenville, Biological effect of ultraviolet radiation on cattle: Bovine ocular squamous cell carcinoma, Am. J. Vet. Res., 40, 783-791, 1979.
Kopecky, K.E., G.W. Pugh, and D.E. Hughes, Wavelength of ultraviolet radiation that enhances onset of critical bovine keratoconjunctivitis. Am. J. Vet. Res., 41, 1412-15. 1980.
Kripke, M.L.. Skin cancer, photoimmunology, and urocanic acid, Photodermatol., 1, 161-163, 1984.
Kripke, M.L., Personal communication, 1989.
Kupfer, C., The conquest of cataract: A global challenge, Trans. Ophthal. Soc. UK, 104, 1-10, 1984.
Ley, R.D., Monodelphis domestica: an animal model for studies in photodermatology including the induction of melanoma, p. 68 in 10th International Congress on Photobiology, Programme and Abstracts, Jerusalem, 1988.
Maitchouk, I.F., Trachoma and cataract: Two WHO targets, Int. Nurs. Rev., 32, 23-25, 1985.
National Cancer Institute (NCI), 1987 Annual Cancer Statistics Review--Including Cancer Trends: 1950-1985, NIH Pub. #88-2789, U.S. Dept. of Health and Human Services, Natl. Inst. of Health, Bethesda, Maryland, 1988.
Noonan, F.P., M.L. Kripke, G.M. Pedersen, and M.I. Greene, Suppression of contact hypersensitivity in mice by ultraviolet irradiation is associated with defective antigen presentation, J. Immunol., 43, 527-533, 1981.
Parrish, J.A.. M.L. Kripke, and W.L. Morison (eds.), Photoimmunology, Plenum Medical Book Co.. 1983.
Perna, J.J., M.L. Mannix, J.E. Rooney, A.L. Notkins. and S.E. Straus, Reactivation of latent herpes simplex virus infection by ultraviolet radiation: A human model, J. Am. Acad. Dermatol., 17. 197-212, 1987.
Pitcher, H.M.. Personal communication, 1989.
Pitts, D.G., A.P. Cullen, and P.D. Hacker, Ocular ultraviolet effects from 295 nm to 365 nm in rabbit eye, Invest. Ophthal. Vis. Sci., 16, 232-240. 1977.
Pitts, D.G., L.L. Cameron, J.G. Jose, S. Lerman, E. Moss, S.D. Varma, S. Zigler, S. Zigman, and J. Zuclich, Optical radiation and cataracts, pp. 5-41 in Optical Radiation and Visual Health, M. Waxler and V.M. Hitchings (eds.), CRC Press, Inc., Boca Raton, Florida, 1986.
Pitts, D.G., The effects of exposure of the eye to ultraviolet radiation, in Environmental Vision (in press), 1989.
Roffo, A.H., Cancer et soleil, Carcinomes et sarcomes provoqués par l'action du soleil in toto, Bull. Frac. Etude Cancer, 23, 590-616, 1934.
Screibner, A., D.E. Hollis, E. Murray, W.H. McCarthy, and G.W. Milton, Effects of exposure to ultraviolet light on epidermal Langerhans cells and melanocytes in Australians of Aboriginal, Asian and Celtic descent, Photodermatol., 3, 15-25, 1987.
Scotto, J., T.R. Fears, and J.F. Fraumeni, Incidence of Non-Melanoma Skin Cancer in the United States, NIH Pub. #82-2433, U.S. Dept. of Health and Human Services, Natl. Inst. of Health, Bethesda, Maryland, 1981.
Slaper, H., Skin Cancer and UV Exposure: Investigations on the Estimation of Risk, Ph.D. Thesis, Utrecht, 1987.
Slaper, H., A.A. Schothorst, and J.C. van der Leun, Risk evaluation of UVB therapy for psoriasis: Comparison of calculated risk for UVB therapy and observed risk in PUVA-treated patients, Photodermatol., 3, 271-283, 1986.
Spruance, S.L., Pathogenesis of herpes simplex labialis: experimental induction of lesion with UV light, J. Clin. Microbiol., 22, 366-368, 1985.
Stem, E., G.A. Mertz, J.D. Stryker, and M. Huppi, Changing animal disease patterns induced by the greenhouse effect. Draft of a preliminary study submitted to EPA, October 1988.
Sterenborg, H.J.C.M. and J.C. van der Leun, Action spectra for tumourigenesis by ultraviolet radiation, pp. 173-190 in Human Exposure to Ultraviolet Radiation: Risks and Regulations, W.F. Passchier and B.F.M. Bosnjakovic (eds.), Elsevier Science Publishers, Amsterdam. 1987.
Takizawa, Y., Some possible ultraviolet effects on the incidence of skin cancer among Japanese due to modifications of the ozone layer, Proc. NIPR Symp. on Polar Meteor. and Glaciol., Natl. Inst. of Polar Res., Tokyo, 1987.
Taylor, D.W. and D.A. Eagles, Assessing the effects of ultraviolet radiation on malarial immunity, draft report to EPA, unpublished, 1989.
Taylor, H.R., Personal communication, 1989.
Taylor, H.R., S.K. West, F.S. Rosenthal, M. Beatriz. H.S. Newland, H. Abbey, and E.A. Emmett, Effect of ultraviolet radiation on cataract formation, New Eng. J. Med., 319, 1429-1433, 1988.
Tucker, M.A., J.A. Shields, P. Hartge, J. Augsburger, R.N. Hoover, J.F. Fraumeni, Sunlight exposure as a risk factor for intraocular melanoma, New Eng. J. Med., 313, 789-792, 1985.
United Nations Development Programme, World Bank. and World Health Organization, Tropical Disease Research, 8th report of the special programme for research and training in tropical diseases (TRD), J. Maurice and M. Pearce (eds.), World Health Organization, Geneva, 1987.
USEPA, Assessing the Risks of Trace Gases that can Modify the Stratosphere, Vol. III, Chap. 6-18, EPA 400/1-87/001C, U.S. Environ. Prot. Agency, Washington, DC, 1987.
USEPA, Ultraviolet Radiation and Melanoma--With a Special Focus on Assessing the Risks of Ozone Depletion, Vol. IV, J.D. Longstreth (ed.), EPA 400/1-87/001D, U.S. Environ. Prot. Agency, Washington, DC, 1987.
USEPA, The Potential Effects of Global Climate Change on the United States, Draft Report to Congress, U.S. Environ. Prot. Agency, Washington, DC, 1989.
Wen-shu, W.M. and Hu Tian-sheng, An epidemiologic survey of senile cataract in China, Chin. Med. J., 95, 813-818, 1982.
Woodhead, A.D., E. Grist, R. Schultz, and R.B. Setlow, A fish model for estimating the wavelengths effective in melanoma induction, p. 32 in 10th International Congress on Photobiology, Programme and Abstracts, Jerusalem, 1988.
{kind=link}
{kind=link}
{kind=link}
{kind=link}