 Reproduced, with permission, from:
Murphy, D. D., and S. B. Weiss. 1992. Effects of climate change on biological diversity in Western North America: Species losses and mechanisms. Chapter 26 in Global Warming and biological diversity, ed. R. L. Peters and T. E. Lovejoy. Castleton, New York: Hamilton Printing.
Reproduced, with permission, from:
Murphy, D. D., and S. B. Weiss. 1992. Effects of climate change on biological diversity in Western North America: Species losses and mechanisms. Chapter 26 in Global Warming and biological diversity, ed. R. L. Peters and T. E. Lovejoy. Castleton, New York: Hamilton Printing.
I. INTRODUCTION
Predictions of the effects of climate change on regional biological diversity are necessarily imprecise. Nonetheless, we must attempt to make projections of the fate of biological systems as the earth warms. Conservation biologists are concerned with the effects of global warming on biodiversity, from extinctions of single populations of highly habitat-specific endangered plants to the extirpation of entire species, communities, and ecosystems. This chapter explores two situations in which reasonably accurate projections do appear possible. First, at a regional scale, we explore the potential effect of global warming on entire montane biotas of mammals, birds, butterflies, and plants in the Great Basin of North America. Second, on a local scale, we examine the potential effects of global warming on the fate of the Bay checkerspot, Euphydryas editha bayensis, a butterfly of native grasslands in the San Francisco Bay area listed as a threatened species by the U.S. Fish and Wildlife Service.
II. CLIMATE CHANGE AND REGIONAL BIODIVERSITY: THE GREAT BASIN
R. L. Peters and J. D. Darling (1985) discuss the potential consequences of global warming on nature reserves, emphasizing both latitudinal and elevational shifts in climate conditions. As climate warms, belts of vegetation on mountain range slopes move upward in elevation. Some high-elevation species may get squeezed out when climate zones exceed the highest elevation in a range or when continuous habitat is reduced to fragments on mountain summits. Since less habitat area exists at higher elevations, extinction probabilities increase for species forced to ascend to track appropriate thermal regimes and resources.
Great Basin mountain ranges are ideally suited for projecting changes in species diversity under such circumstances. The Great Basin of western North America is a region of interior drainages between the Rocky Mountains and the Sierra Nevada. Its arid landscape is characterized by basin and range topography, which has resulted from numerous north-south trending uplifts. These mountain ranges, which rise a thousand or more meters above valley floors, constitute archipelagoes of boreal habitat (fig. 26.1). That boreal habitat consists of riparian plant associations along stream corridors, piñon-juniper forests, subalpine forests, and alpine zones at the highest elevations. The lower boundary of piñon-juniper forest in this region is approximately 2300 m (West et al. 1978), and alpine zones extend from approximately 2750 m to the highest peaks at 3000-4000 m (Billings 1978). Climate projections concur that this region is likely to become both warmer and drier if global warming occurs (see chapter 4).
Boreal habitat was practically continuous in the Great Basin during the Pleistocene. Mountain ranges supported glaciers, basins contained pluvial lakes, and the lower limit of alpine vegetation was approximately 1850 m at glacial maximum (Billings 1978). The distributions of many present-day boreal species were more extensive then. Fossils of the montane vole Phenacomys intermedius, for example, are known from the Toquima Range of central Nevada, far to the southeast of the current distribution of the species. In the same mountain range, fossils of the pika, Ochotona princeps, have been recovered more than 1000 m lower in elevation than the current minimum elevation of the species (Gray- son 1983). Postglacial warming similar to that projected in the immediate future resulted in a 1deg.C rise in temperature from 7500 to 4500 years before the present (Wells 1983). Piñon-juniper forest and timberline apparently shifted upslope by 100-150 m in the White Mountains of-the western Great Basin during that period (Lamarche and Mooney 1967).
A number of studies have established lists of boreal species for major mountain ranges in the Great Basin. Coverage includes mammals (Brown 1971, 1978), birds (Johnson 1978, Behle 1978, Brown 1978), plants (Harper et al. 1978), and butterflies (Murphy and Wilcox 1986, Wilcox et al. 1986, Austin and Murphy 1987). Each of these studies has noted that the distribution of boreal habitat in the Great Basin is analogous to the distribution of habitats across oceanic islands. Several authors have therefore employed island biogeographic theory to explain patterns of species diversity (see MacArthur and Wilson 1967). The essential features of island biogeography pertinent to these studies are: larger islands have more species than smaller islands, islands farther from mainland sources have fewer species than islands of similar size closer to the mainland, and these differences can be explained by opposing rates of colonization and extinction of species.
The results of Great Basin biogeographic studies share several conclusions. All boreal mammal, bird, and butterfly species found in the Great Basin are widely distributed in either one or both of the mainland areas. The major source of less widely distributed species appears to be the Rocky Mountain "mainland" to the east, rather than the Sierra Nevada to the west. Distance from those mainland areas, however, is a poor predictor of species number in most taxa. Furthermore, the flora and fauna of the Great Basin are depauperate compared with those of the Rocky Mountains and Sierra Nevada.
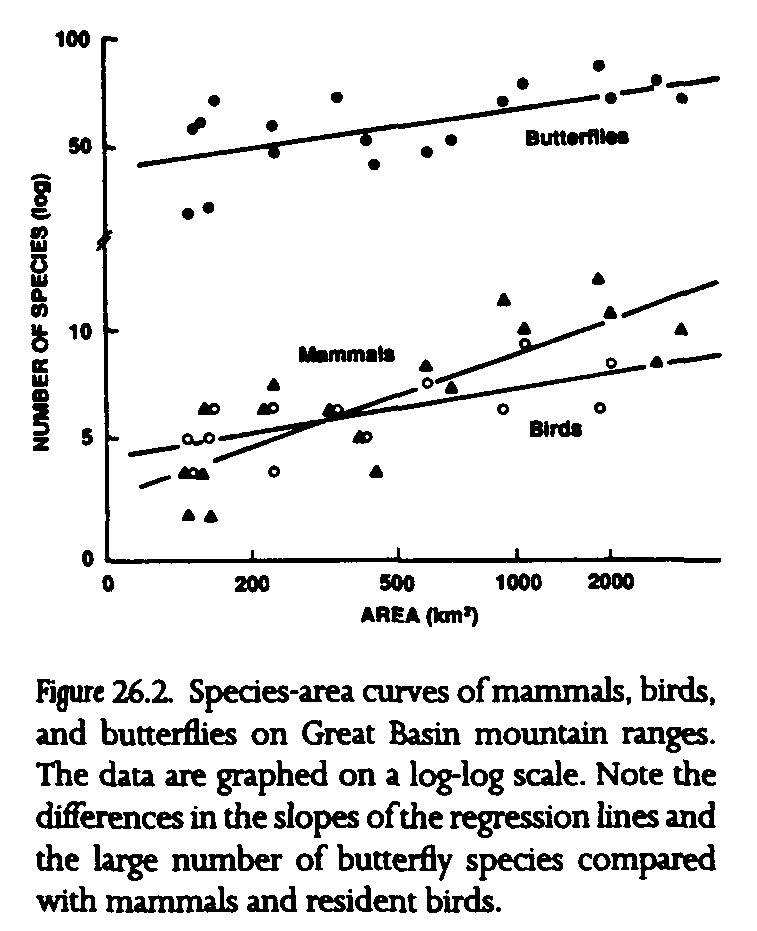
Almost all taxa that have been studied previously exhibit significant increases in species number with increased boreal habitat area (table 26.1, fig. 26.2). Indeed, area explains much of the variance in species numbers for two taxa in particular, 68% in mammals and 84% in plants (see r[2] values in table 26.1), although area explains somewhat less variance in species numbers among ranges for birds and butterflies, at 49% and 39%, respectively. The slopes of the species-area curves also differ among taxa. Species numbers of small mammals decrease relatively rapidly with decreasing area (see z values in table 26.1). Mammal extinctions apparently have not been offset by recolonizations, because nonvolant boreal mammals are largely unable to cross arid lowlands between ranges (Brown 1971, 1978). Mountain ranges lose proportionally fewer species of birds and butterflies with diminishing area, apparently because some recolonization can take place after extinction in these vagile taxa (Brown 1978, Wilcox et al. 1986). The importance of vagility is underscored when butterflies are divided into vagility classes. Sedentary butterflies are more sensitive to decreased area than moderately vagile butterflies. Wide-ranging and migratory butterfly species, in contrast, do not show significant species-area relationships (Wilcox et al. 1986).
The use of area as the fundamental variable in this study should not imply that reduction in habitat area alone is the direct causal factor in extinctions. Many important habitat features contribute to or covary with area of mountain ranges, including elevation of the highest peak, diversity of habitats, and extent of perennial flowing water. Indeed, a habitat diversity score for birds, based on the number of coniferous tree species and the number of major habitat types, has proven to function better than area as a predictor of resident bird diversity (Johnson 1978, Behle 1978, Brown 1978).
Species-area relationships, however, do allow us to generate first-cut estimates of species losses from Great Basin mountain ranges after regional warming of 3deg.C. We make four assumptions in using species-area relationships for these projections:
We are able to estimate whether 10% of present resident species may become extinct in a given mountain range, as opposed to, say, 50% or 80%. Although we would like to differentiate between circumstances in which 30% or 35% of species would be lost, that level of resolution is beyond the predictive capabilities of this model (particularly for ranges supporting already depauperate mammal and resident bird faunas). Some interpretations of species-area statistics have been criticized (e.g., Conner and McCoy 1979), but we believe that the relationships between area of Great Basin mountain ranges and species number in the taxa examined are robust enough to provide a basis for our predictions.
Those caveats stated, we predict that all mountain ranges will suffer reductions and fragmentation of boreal habitat with regional warming of 3deg.C (fig. 26.1). Reductions range from 66% to 90%, with a mean of about 80% (table 26.2). Though a number of central Great Basin ranges currently support more than 900 km[2] of habitat, no range would support more than 400 km[2] of boreal habitat after 3deg.C warming.
Significant numbers of extinctions are predicted in all three animal groups for all mountain ranges. The average loss of mammal species is 44%. Resident birds appear less affected. Butterflies suffer an average decrease of 23% of species per range in this set of mountain ranges. Species losses among sedentary and moderately vagile butterflies are higher, at approximately 30%. The large ranges (Toiyabe-Shoshone, Snake, Schell Creek-Egan, Ruby-East Humboldt, and Monitor-Toquima) have postgreenbouse species numbers comparable to current species numbers in the smaller ranges (Grant-Quinn Canyon and White Pine).
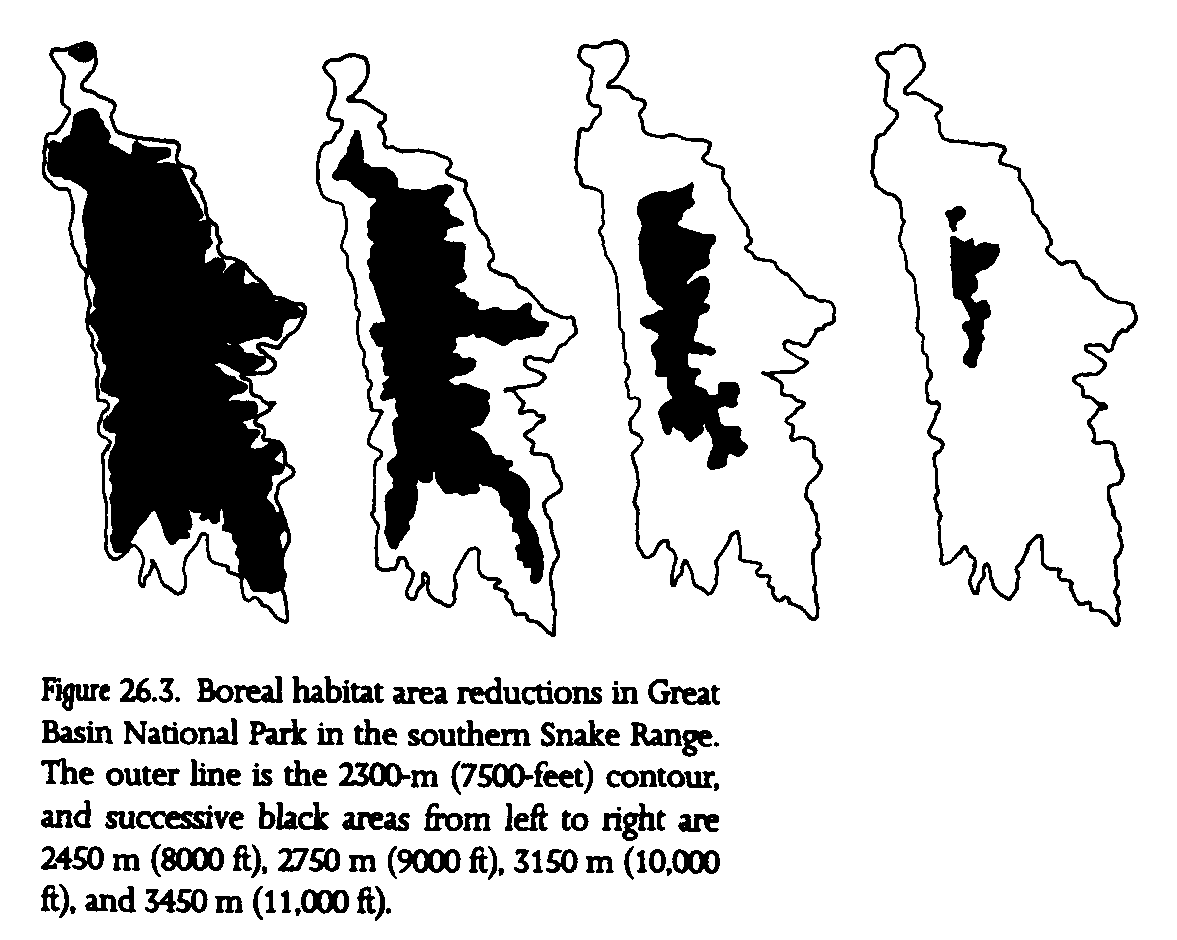
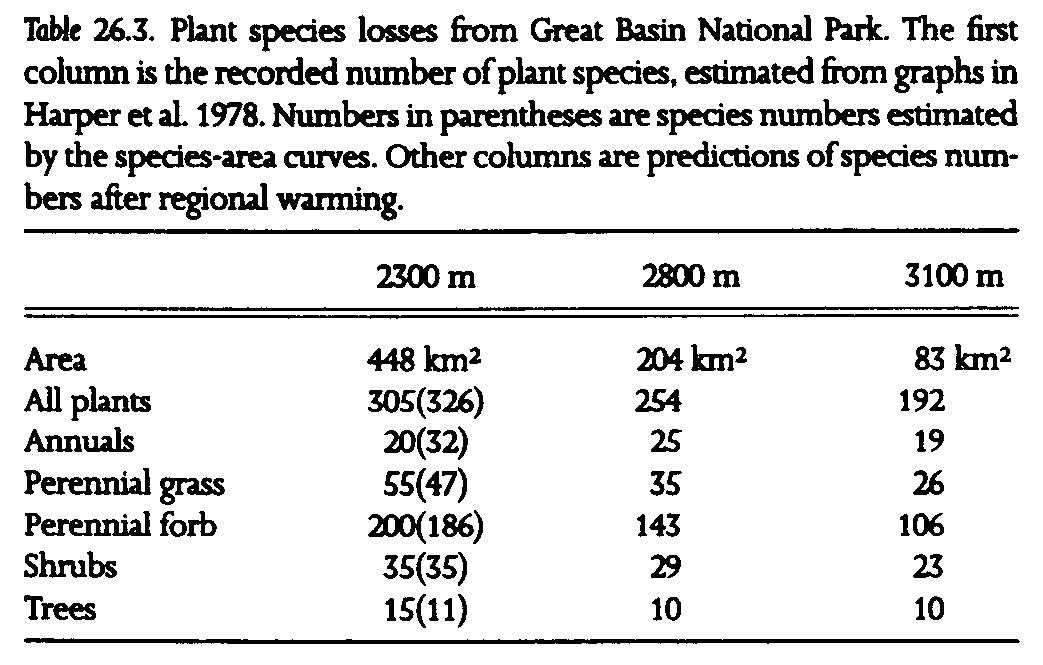
The southern Snake Range was recently designated Great Basin National Park, hence it warrants special note. Previous surveys that included the southern Snake Range established species-area relationships for plants (Harper et al. 1978). We applied these data to the projected reduction of boreal habitat with regional warming in Great Basin National Park (fig. 26.3, table 26.3). Substantial losses of plant species are projected, from 305 currently (326 predicted from the regression equation using fifteen Great Basin ranges) to 254 under the 3deg. warming scenario. Thus, approximately 17% of plant species could be lost from the newest addition to the national park system. A breakdown of plant groups shows that the burden of species losses is not shared equally. Perennial grasses and forbs show the greatest species losses, about 30% (note the high z values in table 26.1), while approximately 17% of shrub species are lost. Great Basin National Park appears to be somewhat buffered against species losses from regional warming, because of its relatively higher proportion of high-elevation habitat to low-elevation habitat, less linear configuration than other ranges, as well as the great height of Wheeler Peak, at nearly 4000 m (fig. 26.1, fig. 26.3).
All in all, these numbers suggest that species losses at the scale of individual mountain ranges will be on the order of 20%-50% after a 3deg.C regional warming, but we cannot say which species will be lost without a more detailed look at the basic biology and habitat requirements of the species. The conjecture that alpine and subalpine zone species will be the likely early victims, since those zones may practically disappear from many mountain ranges, may not necessarily prove correct. Some populations of high-elevation species may actually increase over the short term, should growing seasons extend in length (Ehrlich et al. 1980, Murphy and Weiss 1988c). Although these first-cut approximations tell us that substantial numbers of extinctions are likely to occur, they shed little light on either the time scales or mechanisms of individual species extinctions. In an attempt to identify the factors that could cause local extinction in a 3deg.C global warming scenario, we review here the role of climate in butterfly population dynamics.
III. GLOBAL CLIMATE CHANGE AND THE BAY CHECKERSPOT BUTTERFLY
We now focus on butterflies for a number of reasons. The butterfly extinctions projected above are numerous compared with other animal taxa. Most species require one or at most a few plant species as larval hosts and require an array of adult nectar sources. This dependence on plants usually makes butterflies highly habitat specific. For that reason and others, butterflies as a group are sensitive to thermal conditions and often show immediate dramatic responses to climate fluctuations.
The close relationships of butterflies and their host plants, coupled with the steep species-area curves exhibited by plants (table 26.1), suggest that many butterfly host-plant species may become extinct as habitat area diminishes. Yet. butterfly populations often go extinct well before their plant resources disappear completely, as conditions on both macroclimatic and microclimatic scales become unsuitable.
Many of the complex interactions between macroclimate, microclimate, and population persistence in butterflies are vividly illustrated by long-term studies of checkerspot butterflies in coastal California (see Ehrlich et al. 1975; Ehrlich and Murphy 1981, 1987; Murphy and Weiss 1988a for reviews). Here we discuss how this one species reacts to weather patterns and how habitat topography buffers the effects of weather extremes. These observations form the basis for tentative projections of the fates of butterflies under different local climate change scenarios.
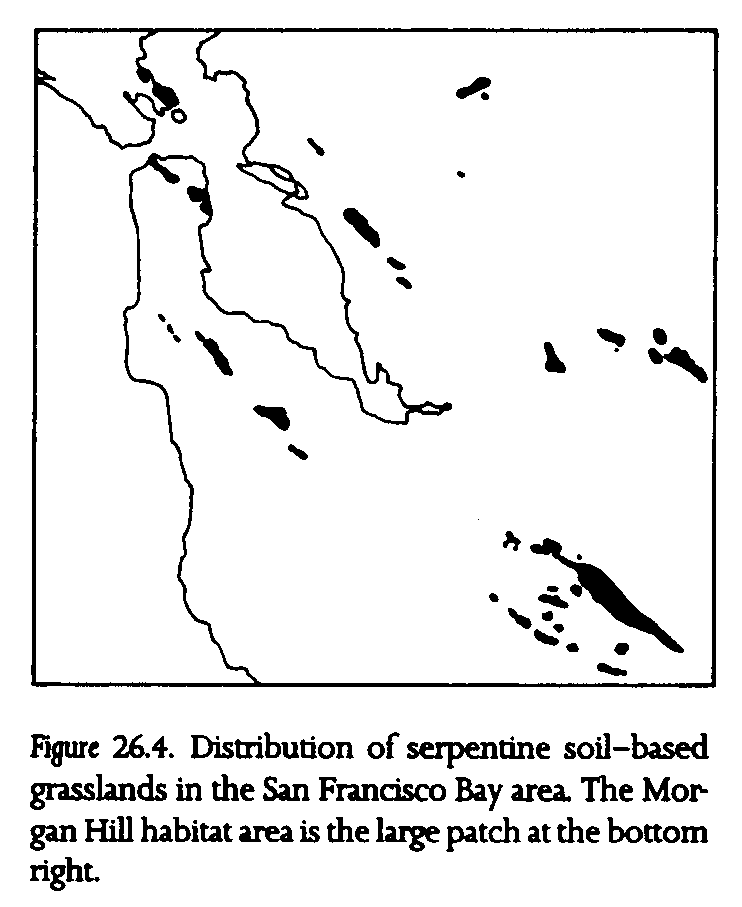
The Bay checkerspot is restricted to patches of serpentine soil-based grassland in the San Francisco Bay area (fig. 26.4). Its recent addition to the federal threatened species list reflects local losses of populations to land development and drought (U.S. Federal Register, Sept. 19, 1987). The serpentine soil-based grasslands inhabited by the butterfly can be viewed as another set of island patches: suitable native grassland habitat in a matrix of unsuitable habitat dominated by alien grassland species.
The life cycle of the Bay checkerspot butterfly is closely tied to the winter-spring growing season of California's Mediterranean climate. Larvae emerge from summer diapause following rains in late fall and early winter. The black larvae feed on the annual forb Plantago erecta and spend much of their time basking in direct sun to speed metabolism and growth (Weiss et al. 1988). Larvae pupate for several weeks, and adults emerge from late February to early May. Females lay egg masses at the base of Plantago and less frequently on the secondary hosts, Orthocarpus densiflorus and O. purpurascens. Eggs hatch after approximately two weeks, and larvae must reach fourth-instar size to enter dry season diapause.
The single most important factor in population size changes in this butterfly is the survivorship rate of larvae as they enter diapause (Singer 1972). Larvae require 4-5 weeks to develop from egg to fourth instar. When winter rains give way to summer drought, the annual host plants of the larvae senesce rapidly. Once Plantago has senesced, larvae may transfer to Orthocarpus, which remains edible later in spring. The temporal phase relationship between adult flight and host-plant senescence and the density of Orthocarpus largely determine subsequent population size, hence the likelihood of population persistence.
This phase relationship is affected by macroclimatic conditions during the winter-spring growing season, suggesting that population persistence may be tied to future changes in regional climate. Sunny winter days speed larval growth and pupal development, and spring rains extend the host-plant growing season. Patterns of rainfall vary widely between years, and extremes of drought and deluge can cause severe population reductions. During droughts, host plants senesce early, and the dry sunny conditions that speed larval growth cannot compensate for early plant senescence (Singer and Ehrlich 1979, Ehrlich et al. 1980). The 1975-77 drought in California caused severe declines in population sizes and numerous extinctions in the southern range of the butterfly in Santa Clara County, including all inhabited patches save the largest in the region (fig. 26.4). The 1987-90 drought period may produce similar results. At the opposite climatic extreme, the wet El Niño weather sequence in 1981-83 also caused population declines because larval and pupal development was slowed markedly by extended rainy weather, and delayed host-plant senescence did not compensate (Dobkin et al. 1987). Weather extremes, in other words, put Bay checkerspot populations at great risk.
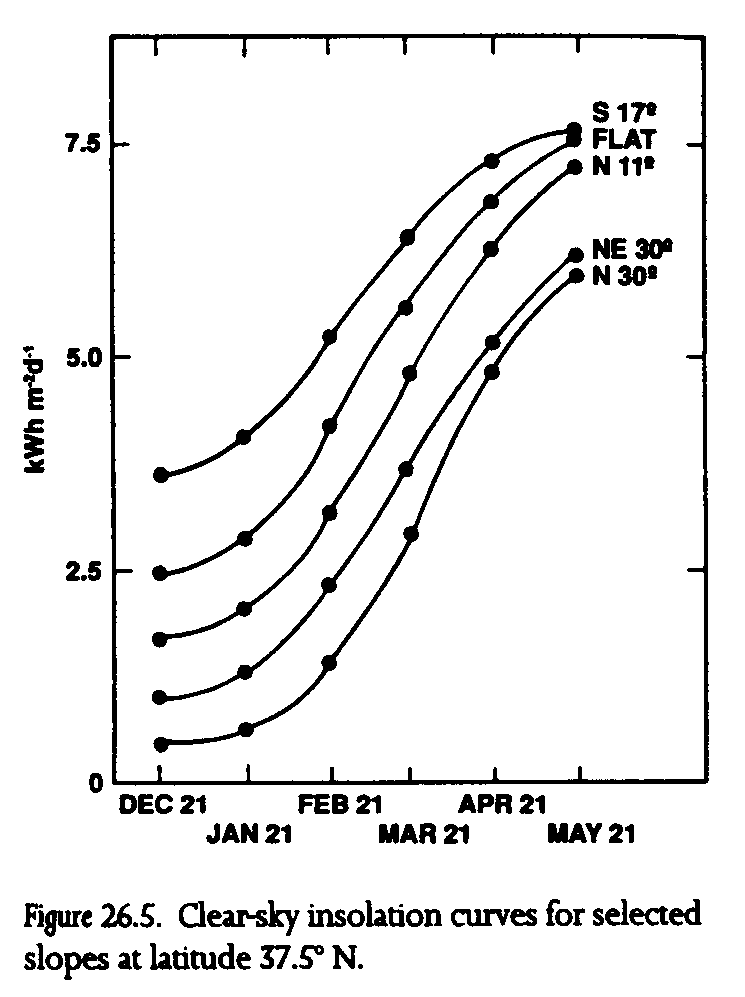
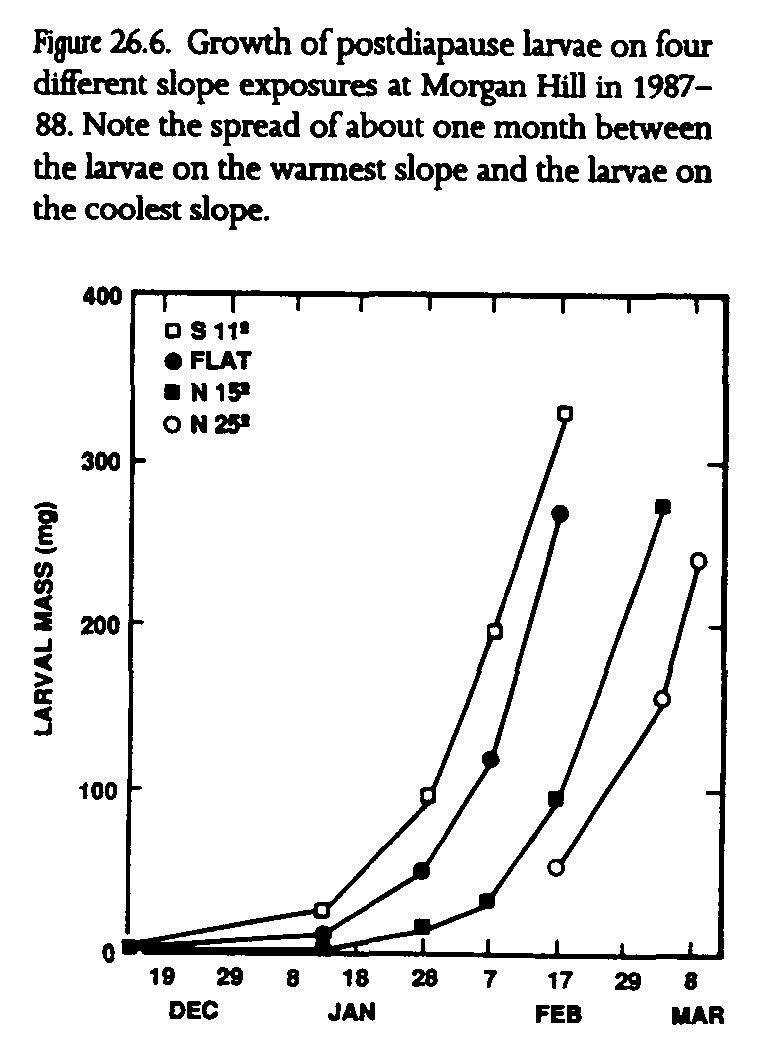
These responses to weather are influenced in each population by the topography of the habitat patch (Singer 1972; Singer and Ehrlich 1979; Weiss et al. 1987, 1988; Murphy and Weiss 1988b). Topographic diversity produces differential solar exposures across habitats, resulting in a complex local mosaic of topographically induced microclimates, or topoclimates (Geiger 1965). Topoclimates can be quantified using calculated clear-sky insolation based on the geometry of sun, earth, and local slope. The shape of insolation curves for different slope exposures is directly reflected in larval development rates and host-plant phenology (fig. 26.5). Larvae on warm south-facing slopes gain an early season growth advantage over larvae on north-facing slopes because south-facing slopes receive significantly more insolation at the winter solstice (at the beginning of post-diapause larval development). Since larval weight gains are directly proportional to insolation, larvae on the warmest habitat slopes may emerge as adults one month earlier or more on average than larvae on the coolest slopes (fig. 26.6). Host plants and adult nectar sources on the warmest slopes grow, flower, and senesce many weeks sooner than those on the coolest slopes. The phase relationship of butterflies and host plants, therefore, varies in space as topoclimates interact with macroclimate (Weiss et al. 1987, 1988; Murphy and Weiss 1988b).
Not surprisingly, spatial patterns of pre-diapause larval survivorship change from year to year, reflecting macroclimatic conditions and the distribution of the previous generation of larvae. Severe droughts tend to shift the larval distribution proportionally toward cooler slopes, which exacerbates the effects of subsequent drought years because few adults are able to emerge early enough for their offspring to find green, edible host plants (Singer and Ehrlich 1979, Ehrlich et al. 1980, Murphy and Weiss 1988b). A shift to warmer slopes during wetter years can buffer a population against subsequent droughts by facilitating earlier adult emergence.
This understanding of the roles of macroclimate and topoclimate in the persistence of Bay checkerspot butterfly populations is crucial to our prediction of the effects of regional warming. More appropriately, we ask whether the Bay checkerspot can survive in its existing habitats, particularly in the large Morgan Hill habitat (fig. 26.4). Because of its size (more than 1500 ha, nearly an order of magnitude larger than any other habitat) and its diverse topography, Morgan Hill supports the population that is most likely to survive a major regional change in climate. In this discussion, the Morgan Hill population may be viewed as existing within the intersection of a suitable habitat space and suitable climate space. The suitable habitat space may be defined as topographically diverse, serpentine soil-based grassland that supports the necessary larval host plants and adult nectar sources. The suitable climate space presently occurs across much of inland San Mateo and Santa Clara counties and the surrounding Inner Coast Range of central California
The Morgan Hill population, however, is in the portion of the distribution that now receives the lowest rainfall because of the rain shadow of the nearby Santa Cruz Mountains. Many populations inhabiting smaller habitat patches adjacent to Morgan Hill were lost during the 1975-77 California drought, while several populations inhabiting smaller patches in wetter San Mateo County survived. For this reason, we believe that optimal mean rainfall for the butterfly may be somewhat greater than that presently occurring at Morgan Hill; hence a macroclimatic shift that results in a moderate increase in rainfall could well enhance the persistence of this species. Conversely, even a slight decrease in mean seasonal rainfall there could result in substantial population declines over time.
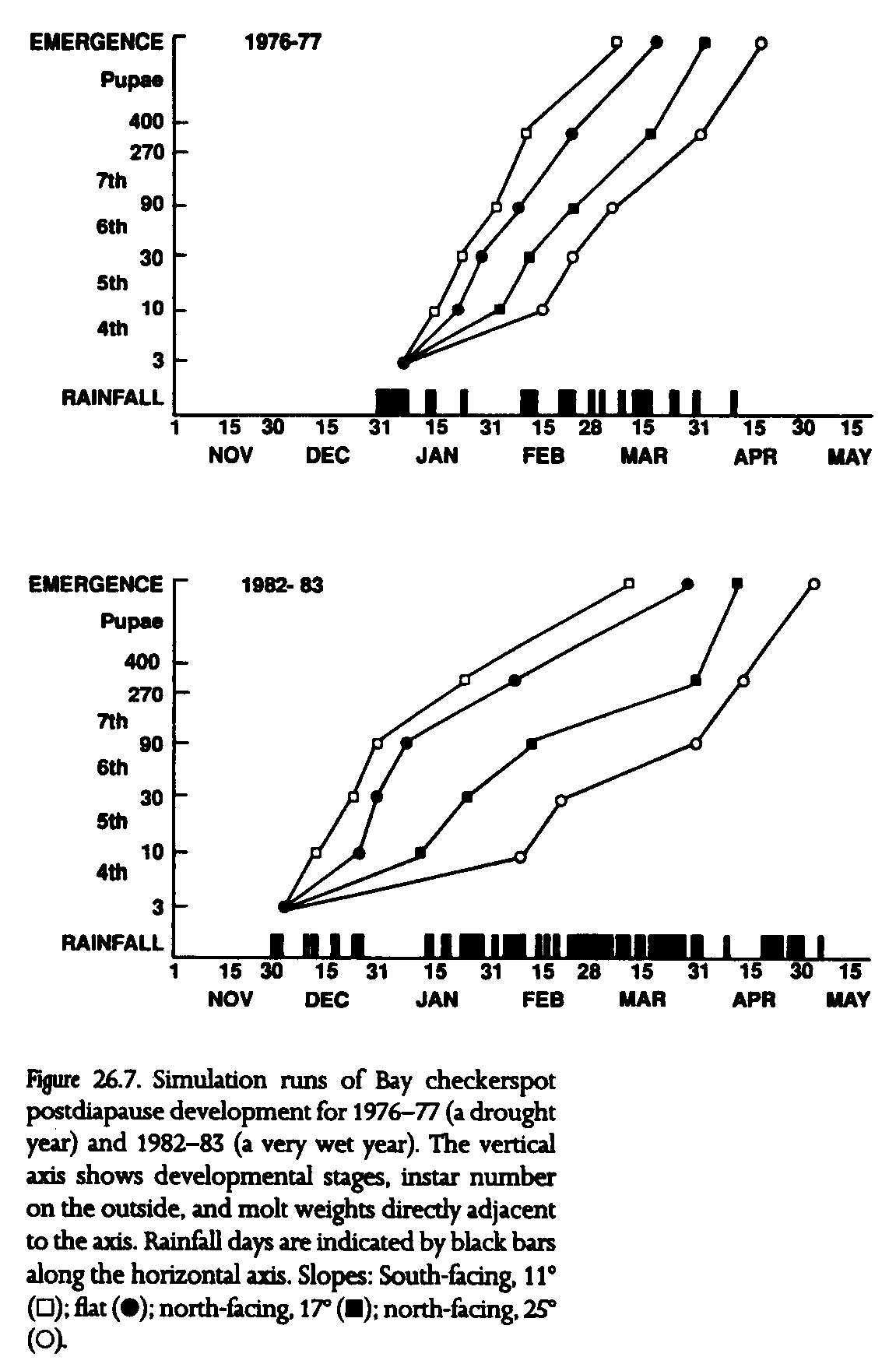
To predict population persistence in an era of climate change, we have developed a simulation model that accounts for the complex interactions among macroclimate, microclimate, and population dynamics of the Bay checkerspot butterfly. We iterate empirical relationships between insolation and growth derived from field studies across an array of slopes over daily weather records for San Jose, near Morgan Hill. The model can predict adult emergence dates from a wide array of topoclimates. We present two examples of that model output (fig. 26.7)--the results of simulation runs for 1976-77, a severe drought year (30% of average annual rainfall), and 1982-83, a very wet year (250% of average).
Gross differences in larval developmental phenologies between these two extreme years include the initiation date of larval feeding, early January in 1976-77 and early December in 1982-83. The larval and pupal periods are much shorter in the drought year (fig. 26.7). Development in much wetter 1982-83 is delayed by extended rainfall in February and March. The mean emergence date (of the four chosen slopes) in 1976-77 is April 1, with a spread of 37 days from the south-facing 11deg. slope to the north-facing 25deg. slope. These figures compare with a mean emergence date of April 7 for 1982-83 and a spread of 52 days. The butterflies that emerged early in the drought year were likely to be the only ones that reproduced successfully, while the early emerging adults in the deluge year were subject to weeks of rain in March and were unable to fly.
Insights gleaned from these and other model runs can be applied to various global warming scenarios for central California. California is located within an unusual climate zone. Temperature increases during an average global warming may be less there than in other regions, and precipitation may increase with global warming because of the proximity of the Pacific Ocean (Kellogg and Schware 1981, Wigley et al. 1981; chapter 20). However, predicting the effects of these changes on the Bay checkerspot butterfly depends on a knowledge of climate specifically during the November-April rainy season at a level of detail as yet unachieved in climate models. Indeed, during overall global warming, coastal California could evidence seasonal cooling. To cover all bases, we tentatively explore four broad cases of local climate change: warmer-drier, warmer-wetter, colder-drier, and colder-wetter conditions.
A warmer and drier climate, similar to that in coastal southern California, would increase the chances of severe drought years, like 1975-76, 1976-77, and 1987-88, which undeniably are detrimental to populations of the Bay checkerspot (Ehrlich et al. 1980, Murphy and Weiss 1988a). Under such circumstances, populations decline when larvae evidence a poor phase relationship with host plant availability. Sequential dry years cause larval populations to become concentrated on cooler slopes (that is, they undergo thermal retreat). Over a sequence of dry years a habitatwide shift of surviving larvae from south-facing slopes to flat areas, flat to north exposures, and north to steeper north exposures occurs, with the result that the warmest exposures cease to contribute significantly to the carrying capacity of the habitat. Populations may become extinct in habitats lacking topoclimatic refugia, particularly in habitats in the driest portions of the distributional range. Emergence in surviving populations is then delayed the following year because most larvae are on cooler slopes, leaving those individuals vulnerable to a subsequent deleterious phase relationship. Most important, an overall net loss of suitable habitat within presently occupied habitats is probable.
A colder and drier climate would be even worse for the Bay checkerspot than a warmer and drier climate. Larval development is slowed by cold weather, even during sunny days. Plant senescence still would occur early because of reduced soil moisture in spring. This combination of extended larval and pupal development with a shortened host-plant growing season is particularly disruptive to the phase relationship between butterflies and their host plants.
Warmer and wetter conditions could result in increases in both sizes and numbers of populations of the Bay checkerspot butterfly. Warm winters in California have long periods of sunny days with daytime highs that produce perfect conditions for rapid development of larvae and pupae. Moisture in such years comes from multiday soaking rains of subtropical origin. Larval and pupal development, therefore, can be relatively rapid, and soil moisture remains high in spring because the ground is saturated late in the season. A good example of such a year is 1985-86, which saw a substantial increase in size of the Morgan Hill population (Murphy and Weiss 1988b). High survivorship because of a favorable phase relationship between larvae and host plants left the majority of larvae residing on warm and moderate slopes--a thermal advance. A sequence of such years could well result in population explosions, reestablishment of peripheral satellite populations, and even defoliation of host plants on slopes heavily populated by larvae. Under such circumstances a population could be a victim of its own success and defoliate the slopes that offer optimal topoclimatic conditions, creating a check on further population growth.
Colder and wetter conditions would make the local climate more like that farther north on the West Coast. Two examples of cold, wet winters are 1981-82 and 1982-83 (El Niño years). This period otherwise produced warm conditions worldwide, pointing to the difficulties of making projections for specific regions (Murphy and Weiss 1988c). Postdiapause larvae at Morgan Hill were largely restricted to cool slopes by 1984 because consecutive spring deluges in 1982 and 1983 delayed larval growth and pupal development more than they extended the host-plant growing season, forcing populations into sequential thermal retreats to cooler slopes. Marked population declines were observed in a number of habitats, although no population extinction events were recorded.
These projections are based on overall trends in climate. Three out of the four scenarios appear unfavorable to the Bay checkerspot, but unpredictable yearly rainfall patterns profoundly affect the impacts of such scenarios on the butterfly. The key factor in determining population persistence is the relative frequency of years causing population declines versus the frequency of years resulting in population increases. Model runs for a variety of years show disparate responses in years with similar rainfall totals, indicating that the timing of rainfall, rather than seasonal totals, is the primary determinant of population fluctuations (Murphy et al. 1990). Furthermore, population responses are dependent on the recent history of the population as manifested by the larval distribution along the topoclimatic gradient.
These projections address only the immediate responses of butterfly populations to climate change. Variations in local temperature and rainfall regimes often result in changes in the composition of plant communities. Such changes can drastically reduce the availability of necessary larval host-plant and nectar resources and can reduce solar exposure at the soil surface. For example, serpentine outcrops in more mesic areas of California tend to be dominated by a chaparral community, with grasslands restricted to the driest, warmest sites. Indeed, the higher elevations at the southern end of the Morgan Hill serpentine outcrop primarily support chaparral species. More moisture could encourage the spread of chaparral species, particularly on the most mesic grassland slopes where isolated shrubs already exist. These north and northeast exposures currently provide a last refuge for the Bay checkerspot in the case of severe multiyear droughts (which are still possible in a wetter climate).
IV. APPLICATIONS OF BAY CHECKERSPOT BUTTERFLY STUDIES TO PROJECTED EXTINCTIONS IN THE GREAT BASIN
How do the Bay checkerspot butterfly studies apply to the projected extinctions in the Great Basin? First, they suggest mechanisms by which climatic factors determine the persistence of populations of butterflies and other herbivorous insects. Our view of the Bay checkerspot butterfly tracking appropriate thermal and moisture regimes from certain slope exposures to others in its grassland habitat assists us in understanding how Great Basin species may track appropriate topoclimates and resources. The process in boreal habitat in the Great Basin can be represented by laterally ascending stripes across mountain slopes: species move both upward and to cooler, more mesic exposures. Common proximate causes of natural local extinctions of butterfiy populations are extreme weather events, particularly season-long drought or deluge and late or early freezes (e.g., Ehrlich et al. 1972). Our studies of the Bay checkerspot indicate that the diversity of topoclimates within a habitat space provides a buffer against such weather patterns. Diverse topoclimates offer potential local refugia for populations faced by erratic and extreme macroclimate conditions. The chance of population extinction, therefore, is reduced in landscapes of high topographic diversity. This observation implies that mountainous regions, such as the Great Basin, may prove to be well buffered against species losses associated with regional warming compared with areas with less relief.
Local topographic diversity, however, can contribute only so much to fending off butterfly extinctions during a prolonged regional warming trend. As demonstrated by our Great Basin habitat projections, some plant species will undergo radical distributional shifts, and others may be unable to disperse rapidly enough to track shifting climate spaces, both circumstances with potentially fatal results for populations of butterflies and other species dependent on them. Many plant distributions will narrow as plants become restricted to higher elevations or to slopes of cooler exposure. Certain topoclimates that now act as refugia in stressful years may become the only available habitat. Conversely, species adapted to xeric conditions may expand their ranges.
All of the factors producing topoclimatic variations apply to Great Basin mountain ranges and provide a framework in which to assess changes on a local scale. The elevational gradient in temperature may be viewed as being fractioned into topoclimates on different slopes. Because they receive more rain, the western sides of these mountain ranges are generally more mesic than the eastern sides. North- and east-facing slopes tend to be more mesic than adjacent south- and west-facing slopes because of lower insolation. Some of these factors have been discussed as they affect species diversity in piñon-juniper forests (West et al. 1978), and numerous other situation-specific examples exist in the literature on mountain ecology. For example, relatively low elevation areas at the base of Great Basin glacial cirques often support alpine species because cold air drainage from higher elevations increases the frequency of frosts (Billings 1978). But, with few exceptions, as species ascend in elevation to follow ascending habitat spaces, they will find that the extents of cool and moist habitats contract.
Even this rather pessimistic view of dwindling habitat spaces is actually optimistic, because it presupposes the ability of organisms to track moving climate spaces and resources. In cases of vagile species like birds or readily dispersed plants, the optimistic view is perhaps justified. But, for the vast majority of species--species that are sedentary, not easily dispersed, highly host-specific, or dependent on other species that have limited vagility---the outlook is grim. Many global warming models suggest a dauntingly short time scale for substantial change, which would not permit populations to make major range adjustments or to evolve in situ to increasing temperature and aridity.
Furthermore, an array of insidious processes is likely to accompany regional warming. For instance, a major mechanism shaping ecosystems in western North America is fire. Lightning fires are a common feature in this largely arid region. One can envision a scenario where parched Great Basin piñon-juniper forests on lower slopes ignite and eliminate mature trees over large areas. Climatic conditions in subsequent years might not allow seedling establishment or survival (see chapter 19 for a more extended discussion of forest regeneration). Many populations of perennial plants depend on intermittent good years for reproduction. Should such years decrease in number, occurring at intervals longer than the lifespans of individual plants or seed bank survival times, then regional extinction could become substantially more likely.
Although we have only begun to address the potential effects of global warming on biological diversity, this chapter and others in this volume echo a common theme---species backed into ecological corners, population disappearances, and disrupted ecosystems. Predicted local and regional climate scenarios promise at the very least a different natural world. The speed at which climate may change assures that this different world will be one that is biologically less rich and less stable than our present one. It will be a world less able to absorb and ameliorate our inevitable future mistakes in land and resource use, a natural world that will, in many ways, seem highly unpredictable and terribly unfamiliar.
Austin, G. T., and D. D. Murphy. 1987. Zoogeography of Great Basin butterflies: Patterns of distribution and differentiation. Great Basin Naturalist 47:186.
Behle, W. H. 1978. Avian biogeography of the Great Basin and Intermountain Region. Great Basin Naturalist Memoirs 2-55.
Billings, W. D. 1978. Alpine phytogeography across the Great Basin. Great Basin Naturalist Memoirs 2:105.
Brown, J. H. 1971. Mammals on mountaintops: Nonequilibrium insular biogeography. Am. Naturalist 105:467.
Brown, J. H. 1978. The theory of insular biogeography and the distribution of boreal birds and mammals. Great Basin Naturalist Memoirs 2-209.
Conner, E. F., and E. D. McCoy. 1979. The statistics and biology of the species-area relationship. Am. Naturalist 113:791.
Dobkin, D. S., I. Olivieri, and P. R. Ehrlich. 1987. Rainfall and the interaction of microclimate with larval resources in the population dynamics of checkerspot butterflies (Euphydryas editha) inhabiting serpentine grasslands. Oecologia 71:161.
Ehrlich, P. R, and D. D. Murphy. 1981. The population biology of checkerspot butterflies (Euphydryas): A review. Biol. Zentral. 100:613.
Ehrlich, P. R., and D. D. Murphy. 1987. Conservation lessons from long-term studies of checkerspot butterflies. Conser. Biol. 22:122
Ehrlich, P. R, D. E. Breedlove, P. E. Brussard, and M. A. Sharp. 1972. Weather and the "regulation" of subalpine butterfly populations. Ecology 53:243.
Ehrlich. P. R, R. R. White, M. C. Singer, S. W. McKechnie, and L. E. Gilbert. 1975. Checkerspot butterflies: A historical perspective. Science 188:221.
Ehrlich, P. R, D. D. Murphy, M. C. Singer, C. B. Sherwood, R. R. White. and I. L Brown. 1980. Extinction, reduction, stability, and increase: The responses of checkerspot butterfly populations to the California drought. Oecologia 46:101.
Geiger, R. 1965. The Climate near the Ground. Cambridge: Harvard University Press.
Grayson, D. K. 1983. Paleontology of Gatecliff Shelter: Small mammals. In The Archeology of Monitor Valley, D. H. Thomas, J. O. Davis, D. K. Grayson, W. N. Melhorn, T. Thomas, and D. Trexler, 2: Gatecliff Shelter. Anthropological Papers of the American Museum of Natural History 59.
Harper, K. T, D. C. Freeman, W. K. Ostler, and L G. Klikoff. 1978. The flora of Great Basin mountain ranges: Diversity, sources, and dispersal ecology. Great Basin Naturalist Memoirs 2:81.
Johnson, N. K. 1978. Patterns of avian geography and speciation in the Intermountain region. Great Basin Naturalist Memoirs 2:137.
Kellogg, W. W., and R Schware. 1981. Climate Change and Society Consequences of Increasing Atmospheric Carbon Dioxide. Boulder, Colo.: Westview Press.
LaMarche, V. C., and H. A. Mooney. 1967. Altithermal timberline advance in western United States. Nature 213:980.
MacArthur, R. H., and E. O. Wilson. 1967. The Theory of Island Biogeography. Princeton: Princeton University Press.
Murphy, D. D., K. E. Freas, and S. B. Weiss. 1990. An "environment-metapopulation" approach to population viability analysis for a threatened invertebrate. Conser. Biol. 4:41.
Murphy, D. D., and S. B. Weiss. 1988a Ecological studies and the conservation of the Bay checkerspot butterfiy, Euphydryas editha bayensis. Biol. Conser. 46:183.
Murphy, D. D., and S. B. Weiss. 1988b. A long-term monitoring plan for a threatened butterfly. Conser. Biol. 2 367.
Murphy, D. D., and S. B. Weiss. 1988c. Drought, deluge, and endangered species. End. Spe. Update 5(6):6.
Murphy, D. D., and B. A. Wilcox. 1986. Butterfly diversity in natural forest fragments: A test of the validity of vertebrate-based management. In Modeling Habitat Relationships of Terrestrial Vertebrates, J. Verner, M. L Morrison, C. J. Ralph, and R H. Barret, eds., pp. 287-292 Madison: University of Wisconsin Press.
Peters, R L, and J. D. Darling. 1985. The greenhouse effect and nature reserves. Bioscience 35:707.
Singer, M. C 1972. Complex components of habitat suitability within a butterfly colony. Science 173:75.
Singer, M. C.. and P. R. Ehrlich. 1979. Population dynamics of the checkerspot butterfly Euphydryas editha. Fortrschr. Zool. 25:53.
Weiss, S. B., R. R. White, D. D. Murphy, and P. R. Ehrlich. 1987. Growth and dispersal of larvae of the checkerspot butterfly, Euphydryas editha. Oikos 50:161.
Weiss, S. B., D. D. Murphy, and R. R. White. 1988. Sun, slope, and butterflies: Topographic determinants of habitat quality for Euphydryas editha bayensis. Ecology 69:1386.
Wells, P. V. 1983. Paleobiogeography of montane islands in the Great Basin since the last galciopluvial. Ecol. Monogr. 53:341.
West, N. E., R. J. Tausch, K. H. Rea, and P. T. Tueller. 1978. Phytogeographical variation within juniper-pinyon woodlands of the Great Basin. Great Basin Naturalist Memoirs 2:119.
Wigley, T. M. L., P. D. Jones, and P. M. Kelley. 1981. Scenario for a warm, high carbon dioxide world. Nature 283:17.
Wilcox, B. A., D. D. Murphy, P. R. Ehrlich, and G. T. Austin. 1986. Insular biogeography of the montane butterfly faunas in the Great Basin: Comparisons with birds and mammals. Oecologia 69:188.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}