 Reproduced, with permission, from:
Balandrin, M. F., J. A. Klocke, E. S. Wurtele, and W. H. Bollinger. 1985. Natural plant chemicals: Sources of industrial and medicinal materials. Science 228: 1154-60.
Reproduced, with permission, from:
Balandrin, M. F., J. A. Klocke, E. S. Wurtele, and W. H. Bollinger. 1985. Natural plant chemicals: Sources of industrial and medicinal materials. Science 228: 1154-60.
Summary. Many higher plants produce economically important organic compounds such as oils, resins, tannins, natural rubber, gums, waxes, dyes, flavors and fragrances, pharmaceuticals, and pesticides. However, most species of higher plants have never been described, much less surveyed for chemical or biologically active constituents, and new sources of commercially valuable materials remain to be discovered. Advances in biotechnology, particularly methods for culturing plant cells and tissues, should provide new means for the commercial processing of even rare plants and the chemicals they produce. These new technologies will extend and enhance the usefulness of plants as renewable resources of valuable chemicals. In the future, biologically active plant-derived chemicals can be expected to play an increasingly significant role in the commercial development of new products for regulating plant growth and for insect and weed control.
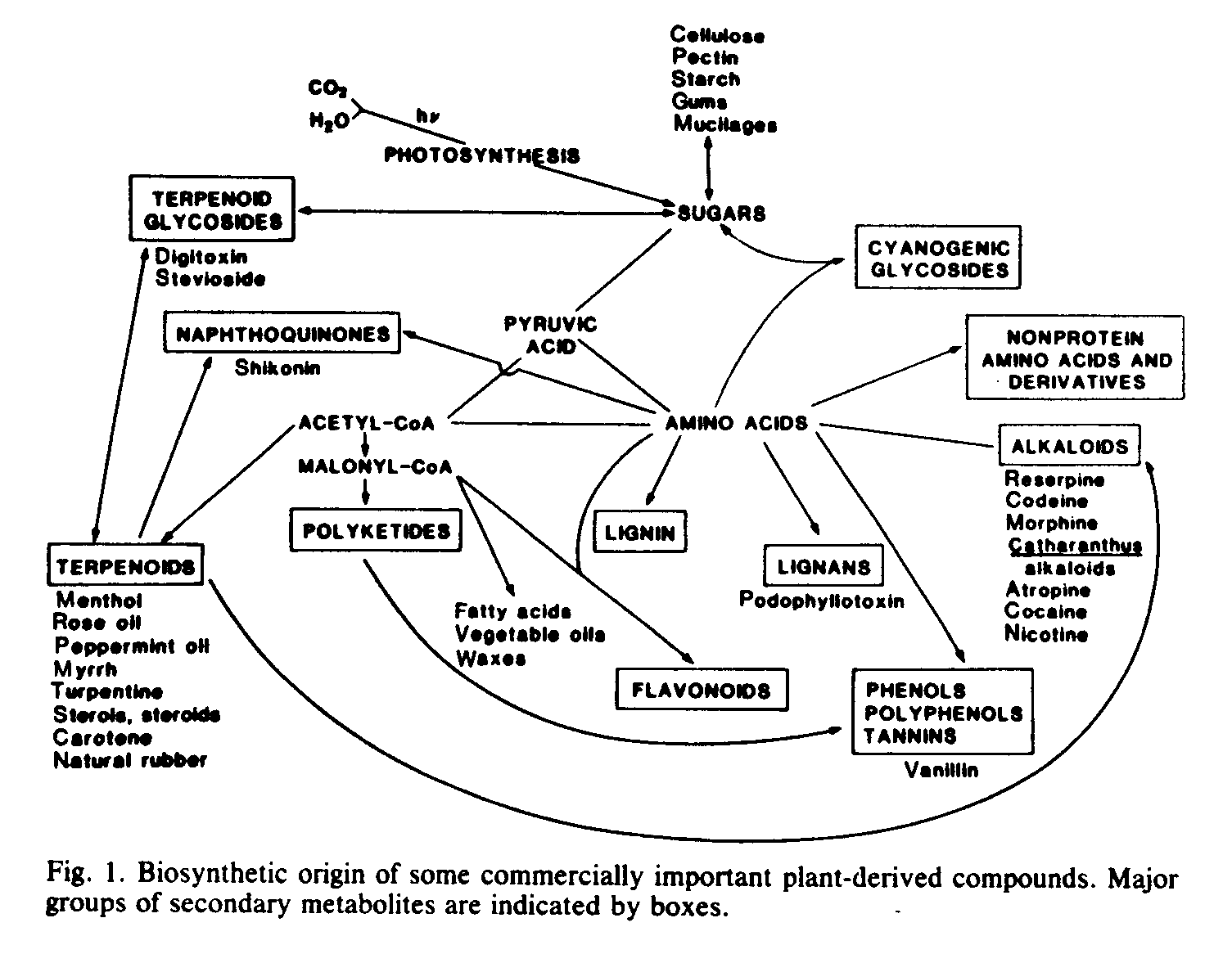
Many higher plants accumulate extractable organic substances in quantities sufficient to be economically useful as chemical feedstocks or raw materials for various scientific, technological, and commercial applications. Natural substances are employed, either directly or indirectly, by a large number of industries, and natural plant products (phytochemicals) figure prominently in several of these (1-3). Economically important plants serve as sources of industrial oils, resins, tannins, saponins, natural rubber, gums, waxes, dyes, pharmaceuticals, and many specialty products (1, 2) (Fig. 1).
For the sake of convenience, plant chemicals are often classified as either primary or secondary metabolites (4, 5) (proteins and nucleic acids are generally excluded from this classification). Primary metabolites are substances widely distributed in nature, occurring in one form or another in virtually all organisms. In higher plants such compounds are often concentrated in seeds and vegetative storage organs and are needed for physiological development because of their role in basic cell metabolism. As a general rule, primary metabolites obtained from higher plants for commercial use are high volume-low value bulk chemicals. They are mainly used as industrial raw materials, foods, or food additives and include products such as vegetable oils, fatty acids (used for making soaps and detergents), and carbohydrates (for example, sucrose, starch, pectin, and cellulose). Such materials are generally valued at less than $1 to $2 per pound ($2.20 to $4.40 per kilogram) and are readily available in large quantities in the marketplace. However, there are exceptions to this rule. For example, myoinositol and ß-carotene are expensive primary metabolites because their extraction, isolation, and purification are difficult.
Secondary metabolites are compounds biosynthetically derived from primary metabolites but more limited in distribution in the plant kingdom, being restricted to a particular taxonomic group (species, genus, family, or closely related group of families). Secondary compounds have no apparent function in a plant's primary metabolism but often have an ecological role; they are pollinator attractants, represent chemical adaptations to environmental stresses, or serve as chemical defenses against microorganisms, insects and higher predators, and even other plants (allelochemics) (5-13). Secondary metabolites are frequently accumulated by plants in smaller quantities than are primary metabolites. In addition, secondary metabolites, in contrast to primary metabolites, tend to be synthesized in specialized cell types and at distinct developmental stages, making their extraction and purification difficult. As a result, secondary metabolites that are used commercially as biologically active compounds (pharmaceuticals, flavors, fragrances, and pesticides) are generally higher value-lower volume products than the primary metabolites. Thus, compared to primary metabolites (bulk chemicals), many secondary metabolites can be considered as specialty materials or fine chemicals.
Examples of commercially useful plant secondary metabolites are nicotine, the pyrethrins, and rotenone, which are used in limited quantities as pesticides (10), and certain steroids and alkaloids, which are used in drug manufacturing by the pharmaceutical industry (1). The steroids and alkaloids include steroidal sapogenins, Digitalis glycosides, the anticancer Catharanthus (formerly Vinca) alkaloids, belladonna alkaloids (for example, atropine, hyoscyamine, and scopolamine), cocaine, colchicine, opium alkaloids (codeine, morphine, and papaverine), physostigmine, pilocarpine, quinine, quinidine, reserpine, and d-tubocurarine (1, 14-19). Other secondary plant metabolites are used in limited quantities as pharmacological tools to study various biochemical processes. For example, diterpene esters (among which are phorbol derivatives) from the latices of various species of Euphorbia (members of the spurge family) are potent irritants and cocarcinogens and are useful in studies of chemical carcinogenesis (20).
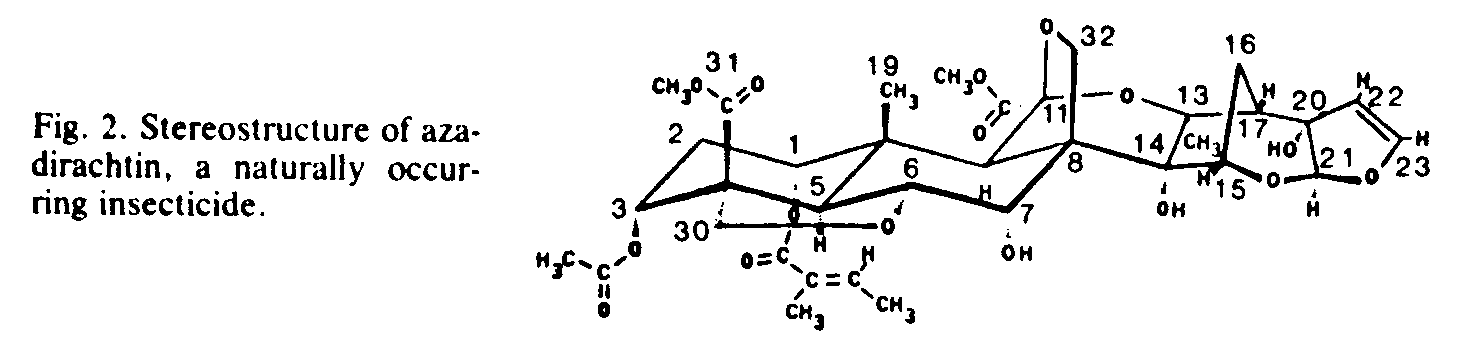
Compared to the relatively low cost of primary or bulk metabolites, secondary plant metabolites are often valued at several dollars to several thousand dollars per pound. For example, purified opium alkaloids (codeine and morphine) are valued in the range of $400 to $600 per pound ($650 to $1250 per kilogram), while rare volatile (essential) oils such as rose oil are often valued at over $1000 per pound ($2000 to $3000 per kilogram). Phorbol esters have a retail value of about $2000 per gram. The anticancer Catharanthus alkaloids have a wholesale value of about $5000 per gram, and their retail value may be as much as $20,000 per gram (21). Secondary natural products often have highly complex structures with many chiral centers. which may determine biological activity; such complex compounds cannot be synthesized economically (16, 18, 22). A good example of a secondary metabolite having a high degree of structural complexity is the naturally occurring plant insecticide azadirachtin (Fig. 2) (22).
Economically important primary and secondary metabolites share several common characteristics. Most can be obtained from plant materials by steam distillation or by extraction with organic or aqueous solvents, and (excluding the biopolymers, natural rubber, condensed tannins, and the high molecular weight polysaccharides such as gums, pectin, and starch) they tend to be relatively low in molecular weight (generally less than 2000).
Few purified plant proteins are used commercially. Economically important plant-derived enzymes include papain and chymopapain [enzymes derived from papaya (Carica papaya) that are used medicinally and as meat tenderizers], bromelain (a protein-digesting and milk-clotting enzyme from pineapple juice), and malt extract (a product from barley that contains amylolytic enzymes) (1).
The extraction and use of specialized plant proteins from plant cells has limited importance for several reasons. First, their chemical structure imposes certain constraints on their use as biologically active compounds, that is, as drugs and pesticides. For example, most proteins cannot be absorbed readily through mammalian skin or insect exoskeleta, and most also cannot be administered orally (except to achieve local effect) because they are subject to digestive degradation by proteolytic enzymes. For reproducible systemic effects to be elicited, most polypeptides (such as chymopapain) must be administered by injection. Thus, proteins are not made biologically available as easily as are secondary metabolites (protein products), thereby complicating product formulation and use. For example, certain potentially useful pesticidal proteins may be rapidly degraded in the field because of physicochemical instabilities. Second, the technology for the insertion and expression in bacteria and yeast of the genes coding for valuable polypeptides already exists. This obviates the need to grow plant cells to obtain protein products since genetically engineered microorganisms can be grown efficiently in culture. However, genetic engineering of bacteria or yeast to produce the complex secondary metabolites naturally biosynthesized by certain plant species is more difficult because of the nature of secondary metabolite biosynthesis. Proteins are immediate gene products, while secondary metabolites are usually biosynthesized via the joint action of many gene products (enzymes) (23). Genes responsible for the biosynthesis of economically important secondary metabolites have not yet been isolated (for each biosynthetic pathway leading to the production of a secondary metabolite, many genes are required): furthermore, microorganisms have many enzymes that may catalyze unwanted side reactions with the desired metabolite or intermediates in its biosynthetic pathway. Thus, at least in the immediate future, plants or plant cells will probably continue to serve as the sources for most of these plant-specific materials.
Secondary Metabolites in Foods, Beverages, and Pharmaceuticals
Most spices, condiments, teas, and other beverages such as coffee and cocoa owe their individual properties (flavors and aromas) to the pharmacologically active secondary plant metabolites that they contain. Although a number of these active substances (such as vanillin, ephedrine, and caffeine) are produced by semi- or total synthesis, high prices are still paid in some cases for compounds extracted from their natural sources, especially if they are intended for use as food additives or flavoring agents.
Some biologically active secondary metabolites have found application as drug entities or as model compounds for drug syntheses and semisyntheses (1, 14-19, 24). However, it is frequently forgotten that natural products often serve as chemical models for the design and total synthesis of new drug entities (24, 25). For example, meperidine (Demerol), pentazocine (Talwin), and propoxyphene (Darvon) are all totally synthetic analgesic drugs for which opiates such as morphine and codeine were the models, while aspirin is a simple derivative of the naturally occurring salicylic acid originally derived from willows (Salix spp.).
The commercial value of drug products derived from higher plants should not be underestimated. For example, in 1980 American consumers paid about $8 billion for prescription drugs derived solely from higher plants. From 1959 to 1980, drugs derived from higher plants represented a constant 25 percent of all new and refilled prescriptions dispensed from community pharmacies in the United States (this does not take into account nonprescription drug products or drugs used exclusively in hospitals) (15-19).
Plants continue to be important sources of new drugs, as evidenced by the approvals in the United States in 1983 of two new plant-derived drugs. Etoposide, a semisynthetic antineoplastic agent derived from the mayapple (Podophyllum peltatum), is reported to be useful in the chemotherapeutic treatment of refractory testicular carcinomas, small cell lung carcinomas, nonlymphocytic leukemias, and non-Hodgkin's lymphomas (26). Atracurium besylate, a skeletal muscle relaxant, is another new plant-based drug approved for use in the United States; it is structurally and pharmacologically related to the curare alkaloids (26). Important plant-derived drugs that are still obtained commercially by extraction from their whole plant sources are listed in Table 1.
Pesticides, Allelochemicals, and Plant Growth Regulators
Pesticides. Extracts of plants have been used as insecticides by humans since before the time of the ancient Romans, a practice that continues to the present with many of the 2000 species of plants known to have insecticidal properties (27). The use of "insecticidal" plants is especially prevalent in the developing countries, where plants grown locally (which are renewable resources) are cheaper for subsistence farmers to use than are synthetic chemical pesticides (which are petrochemical derivatives). Commercially, however, only a few of these plants---those containing pyrethrins, rotenoids, and alkaloids---have been used to any extent in the United States as sources of insecticides (10, 28).
The most economically important of the natural plant compounds used in commercial insect control are the pyrethrins, a group of six closely related esters extracted from pyrethrum flowers. Pyrethrum has been used as an insecticide since at least the early 1800's in Persia and Yugoslavia. By 1828 pyrethrum was being processed for commercial insect control, and by 1939 imports of pyrethrum to the United States reached a peak of 13.5 million pounds. Use of the natural product declined in the early 1950's because of the advent of synthetic analogs (for example. allethrins), which were both more stable and more effective in the field. Nevertheless, when properly formulated with antioxidants or stabilizers (including the natural plant compounds tannic acid and hydroquinone) and synergists (including the natural plant compounds sesamin and myristicin), the pyrethrins are still economically viable insecticides. In fact, the demand worldwide for the pyrethrum flowers remains in excess of 25,000 tons annually and is satisfied by the estimated 150 million flowers still hand-harvested daily in Kenya, Tanzania, and Ecuador (29).
Rotenone and the rotenoids have long been used as insecticides and piscicides. By the early 1950's more than 7 million pounds of Leguminosae roots (Derris, Lonchocarpus, and Tephrosia spp.) containing these insecticides were imported annually to the United States. In 1972, about 1.5 million pounds were used in the United States for pest control in the home and garden markets and to control ectoparasites on animals (30). These compounds have also been used to control fish populations. For example, the Brazilian government used thousands of tons of the piscicidal root dust containing rotenoids in the specific removal of piranhas from Brazilian rivers and reservoir systems (31).
Among the most important of the natural alkaloids used in insect control have been nicotine and the related compound nornicotine. The use of these insecticidal alkaloids dates back to the 1600's and grew to 5 million pounds by the mid-1900's. Since then, the annual worldwide production of nicotine has dropped to about 1,250,000 pounds of nicotine sulfate and 150,000 pounds of nicotine alkaloid because of the high cost of production, disagreeable odor, extreme toxicity to mammals, and limited insecticidal activity (30, 32). The structurally related compound anabasine (neonicotine) is currently in commercial use in the Soviet Union (10). Other less important insecticidal alkaloids include veratrine [a mixture of alkaloids (cevadine, veratridine, and sabacilline)] and ryanodine. Physostigmine, another alkaloid that is isolated from the calabar bean (Physostigma venenosum), served as a model compound for the development of the carbamate insecticides (33).
Allelochemicals. Allelochemicals are substances produced by higher plants that selectively inhibit the growth of soil microorganisms or other plants (or both). These phytotoxic compounds play a role in chemical warfare between plants (allelopathic interactions) and include natural herbicides, phytoalexins (microbial inhibitors), and inhibitors of seed germination. Although many allelochemicals are strictly defense substances, others are offensive compounds that act directly in weed aggressiveness, competition, and the regulation of plant density. Allelopathic agents encompass a wide array of chemical types, including volatile terpenoids. phenylpropanoids, quinones, coumarins, flavonoids, tannins and other phenolics, and cyanogenic glycosides. Although none of these compounds is currently used commercially, there are expectations that some may provide models for new synthetic or semisynthetic herbicides and antimicrobials (6, 11, 13).
Plant growth regulators. The compound brassinolide represents a new generation of natural plant growth regulators with possible commercial utility in agriculture (34, 35). A steroidal lactone originally isolated from the pollen of rape (Brassica napus), brassinolide, can promote and accelerate plant growth at low concentrations (nanograms per plant) (35). This compound and its semisynthetic analogs (the brassinosteroids) may increase plant biomass by promoting both cell expansion and cell division, unlike the agriculturally important gibberellins, which stimulate cell expansion (34, 35). In field tests, brassinosteroids increased crop yields, depending on the particular crop and environmental conditions (34). Brassinosteroids could be produced commercially by semisynthesis from cheaper and more plentiful naturally occurring steroid starting materials in a manner analogous to the commercial production of pharmaceutically important steroid hormones from plant-derived steroid sapogenins. It has been estimated that 1 gram of a semisynthetic brassinosteroid could be produced commercially for $5 to $10. Since these compounds exert their growth-promoting activity at low concentrations (1 part per million in solutions used in field tests), up to 5 acres could be treated with this quantity of material.
Prospects for Discovering New Bioactive Compounds from Plants
It has been estimated that only 5 to 15 percent of the 250,000 to 750,000 existing species of higher plants has been surveyed for biologically active compounds (16, 18, 36). Such estimates can be misleading, however, since it is often the case that investigated plants have been screened for a single type (or, at best, a few types) of biological activity (16). The best example of an extensive but narrow screening program is the National Cancer Institute's search for antitumor agents from higher plants (37). Over a 20-year period (1960 to 1981), some 120,000 plant extracts representing approximately 35,000 species were tested solely for cytotoxic or antitumor activity (16, 37). During the screening, natural compounds useful as new drugs for other ailments or conditions (for example, analgesics, antiarthritics, and antipsychotics) and substances exerting insecticidal or allelopathic effects were undoubtedly overlooked. Thus it appears that the plant kingdom has received little attention as a resource of potentially useful bioactive compounds. Since many secondary metabolites are genus- or species-specific, the chances are good to excellent that many plant constituents with potentially useful biological properties remain undiscovered, undeveloped, and unused.
In spite of the advances in extraction technology, separation science (chromatographic techniques), and analytical and spectroscopic instrumentation, we still know little about the secondary metabolism of most of the higher plant species. This is especially true in the case of tropical floras. Although the tropics contain most of the world's plant species, it has been estimated that more than half of these are unknown and that most have never been surveyed for chemical constituents (38, 39). For example, it has been estimated that nothing is known about the chemistry of more than 99 percent of the plant species comprised by the flora of Brazil (38). This is alarming in view of the current rate of extinction of tropical floras, especially forests, before their plants have been adequately cataloged and studied. If the current trend of destruction of tropical forest habitats continues at its present rate, phytochemists may have only a few decades remaining in which to survey the chemical constituents of a large part of the plant kingdom for potentially useful compounds (39).
The development of new medicinals from higher plants is costly and time-consuming because those readily available plants having pronounced pharmacological effects (such as the opium poppy, foxglove, and belladonna) have been known for centuries and have been thoroughly investigated (40). However, the prospects for developing new pesticides and herbicides from plant sources (allelochemicals) are better because this area of investigation is newer than the area of medicinal plant research. Unfortunately, the current trends in the destruction of tropical forests will eliminate many significant opportunities for research and development in these areas.
Natural plant chemicals will undoubtedly play a significant role in the future of pest control in both industrialized and developing countries. Because of the need for new, safer insecticides, there have been efforts to exploit the natural chemical defense mechanisms of plants (biorational approach and design) (41). Much of the research has focused on plant chemicals that specifically affect processes peculiar to the target pests-- for example, agents that affect insect feeding behavior (feeding deterrents) and insect growth regulators, which specifically disrupt the endocrinological balance of pest insects.
Many plant chemicals deter insects from feeding (8, 42). Although none of these chemicals (antifeedants) have yet been developed commercially for that purpose, some have that potential. For example, azadirachtin (Fig. 2) and other limonoids in many plant species in the Meliaceae and the Rutaceae have long been used with success in insect control, especially in India (43). A number of chemical companies are evaluating (and in some cases developing) plant extracts containing limonoids for commercial insect control. The complex structures of the limonoids preclude their economical chemical synthesis, but their availability from plant sources may warrant commercialization. For example, azadirachtin (Fig. 2) is available in the estimated 14 million Azadirachta indica trees in India and the countless Melia azedarach trees worldwide (44). Other antifeedant limonoids are available as industrial byproducts, including limonin, nomilin, and obacunone from the citrus industry and gedunin from the timber industry (45).
Insect growth regulators, including analogs and antagonists of endogenous hormones, have also been identified in plants. Prominent among these are the analogs of two insect hormones (molting hormone and juvenile hormone) and the antagonist for juvenile hormone. Chemicals structurally similar or identical to the insect molting hormone have been found in many plants, especially ferns and yews. However, the complexity of the steroid nucleus and the weak insecticidal effect when applied topically or orally to most insects preclude the commercial development of these plant compounds as insecticides. The only commercial use of the molting hormone analogs has been in the sericultural industry for the synchronization of cocoon spinning of silkworm colonies (46).
Analogs of the insect juvenile hormones have also been found in plants, including the juvocimenes in Ocimum basilicum, juvabione in Abies balsamea, and farnesol in many plant oils. These natural plant products have never been used commercially, but they have served as model compounds for the development of juvenile hormone analogs (for example, methoprene and kinoprene) that are available for the control of mosquitoes, flies, fleas, and some stored-products pests (47).
Chromenes have been isolated from Ageratum houstonianum as juvenile hormone antagonists (48). Thus far, however, chromenes and their analogs have been found to be effective on relatively few species of insects, and their potential for commercialization may therefore be limited.
Other plant species that contain secondary metabolites with potential use as commercial insecticides include Acorus calamus, Artemisia tridentata, Heliopsis longipes, Mammea americana, and Tagetes minuta (3, 10).
Cell Suspension Cultures as Alternative Sources of Plant Natural Products
As the natural habitats for wild plants disappear and environmental and geopolitical instabilities make it difficult to acquire plant-derived chemicals, it may become critical to develop alternative sources for important natural plant products. There has been considerable interest in plant cell cultures as a potential alternative to traditional agriculture for the industrial production of secondary plant metabolites (49, 50). This interest has given rise to considerable research in Japan, West Germany, and Canada (21).
Cell suspension systems could be used for the large-scale culturing of plant cells from which secondary metabolites can be extracted. The advantage of this method is that it can ultimately provide a continuous, reliable source of natural products. In addition, compounds from tissue cultures are more easily purified because of the absence of significant amounts of pigments, thus reducing production costs. The basic technologies for obtaining high yields of specific secondary metabolites from large-scale cultures are still being developed (21, 51, 52).
In plants, high concentrations of secondary metabolites tend to accumulate in specific cell types at specific developmental stages. Thus, it is not surprising that tissue-cultured cells from these plants typically accumulate large amounts of secondary compounds only under specific conditions of cell culture (52). Maximizing the production and accumulation of secondary metabolites by tissue-cultured cells requires (i) selecting proper genotypes, (ii) selecting high-yielding cell clones, and (iii) manipulating the parameters of the environment and medium (52, 53). In this way more than 30 natural products have been synthesized in cell culture in amounts larger than those produced in whole plants (54).
Selection of proper genotypes. Cell cultures derived from individual whole plants that accumulate large amounts of a secondary metabolite also tend to accumulate larger amounts of that metabolite (52, 53, 55, 56). This has been shown in comparisons of the concentrations of harmane alkaloid and serotonin in seeds, seedlings, and callus cultures of Peganum harmala (56).
Selection of high-yielding cell clones. Plant cells undergo continuous process of epigenetic or genetic changes during culture (53, 57). Therefore, an ongoing selection for cell clones or lines that are high producers of the desired chemical compound may be required (53, 55). Such a strategy resulted in a 55-fold increase in a 12ß-hydroxylation reaction in a 20-liter fermenter culture over a 4-year period (58).
Manipulation of parameters of the environment and medium. Alterations in hormones and other media components affect the rates of both cell growth and accumulation of secondary metabolites (52). Great increases in yield have been obtained in systematic studies that were based on what is known about the biosynthetic pathways, physiology, and localization of secondary metabolites in plants (51-53). In the intact plant, synthesis of metabolites thought to be protective compounds is induced by fungal infection. The addition of fungus or compounds produced upon fungal infection (elicitors) that mediate the synthesis of secondary metabolites has resulted in hemigossypol formation by cultures of Gossypium arboreum cells (59). Cells of poppy plants (latex cells), which synthesize opium alkaloids, have a distinctive morphology. By selecting for cells with a similar morphology in culture, opium alkaloid synthesis was increased (60). In one of the most comprehensive studies of the effects of physicochemical factors on secondary metabolite biosyntheses, the effects of ultraviolet light, culture age, and fungal elicitors on the accumulation of flavonoids in parsley cell cultures have been examined with regard to biosynthetic enzyme activities, enzymology, synthesis of messenger RNA, and the functioning of related biosynthetic pathways (61).
Partially because of the complexity of secondary metabolite biosynthesis, the enzymology, regulation, cellular and subcellular sites of biosynthesis (as opposed to sites of accumulation), and even the biosynthetic pathways are poorly understood for many compounds. It may well be that elucidation of the physiological and biochemical factors governing secondary metabolite biosynthesis and degradation will contribute to our ability to increase amounts of secondary products in cultured plant cells. For example, acetyl-CoA (coenzyme A) carboxylase catalyzes the formation of malonyl-CoA, an intermediate required for the biosynthesis of many natural products (Fig. 1). Studies indicate that acetyl-CoA carboxylase is compartmentalized at the cellular and subcellular levels, that it may have tissue-specific isozymal forms, and, in at least one instance, that its activity increases as flavonoid biosynthesis is induced (61, 62). However, despite its central role in many biosynthetic pathways, little more is known about the role of acetyl-CoA carboxylase in regulating the biosynthesis of secondary metabolites. In another example, although high rates of degradation and turnover of many secondary metabolites have been shown in plants, the implications of turnover in attaining high rates of secondary metabolite production in cultured cells have been only rarely examined (63).
Facilities for the large-scale production of secondary metabolites from plant cells have been developed, but further research is needed to improve their economic efficiency. Briefly, production methods include (i) fermentation vats similar to those used for the commercial culturing of microorganisms but with gentler mixing procedures to avoid disruption of plant cells and (ii) immobilized continuous cultures (51, 52). The former method offers the advantage that batch cultures can be used to harvest nonexcreted metabolites. The latter resolves problems of cell damage during mixing, enables easy and continuous change of media and harvest of (excreted) secondary metabolites, and negates the disadvantages of the slow growth of plant cells and their tendency to produce most secondary products in a differentiated (specialized) state (52).
An alternative method of secondary metabolite production is the use of plant cell culture for the conversion of exogenous precursors to desired metabolites (biotransformation). An example is the stereospecific 12ß-hydroxylation of exogenous ß-methyldigitoxin to ß-methyldigoxin by Digitalis lanata cells. By making appropriate selections of clones and manipulations to their environment, the rate of this biotransformation was increased by 60-fold over several years (58). Biotransformation has additional potential as a means of achieving the biosynthesis of novel compounds by supplying to the plant cells precursor compounds that are not biosynthesized by the plant itself (51, 53). However, in the future, simple biotransformation reactions may be carried out by immobilized enzymes. Alternatively, biotransformation reactions involving only one or two genes are potential candidates for genetic engineering of the relevant genes into easily cultured microorganisms.
Cost analyses indicate that by means of current technology, production of a secondary metabolite in plant cell culture is economical for cultures producing more than 1 gram of compound per liter of cell culture for compounds with a value exceeding $500 to $1000 per kilogram (21, 64). The total market for the compound must be sufficiently large to warrant the capital expenditures needed to develop a tissue culture system (21). In view of such economic constraints candidates for commercial production by plant tissue culture are limited to a few types of high-value, plant-specific compounds. These include diosgenin-derived steroid hormone precursors, Digitalis glycosides, opium alkaloids (codeine and morphine), the Catharanthus alkaloids, and, possibly, complex mixtures such as essential oils (for example, rose oil). Five to twenty times more Catharanthus alkaloids (potential anticancer agents) were obtained by tissue culture methods than from whole plants (21). Accumulation of Catharanthus alkaloids at about 75 percent of the accumulation from small-scale cultures (0.07 liter) has been achieved from cells grown in a 750-liter production fermenter (51). A German pharmaceutical company is going ahead with plans to produce Digitalis glycosides in tissue culture systems within the next few years (21). Mitsui Petrochemical Industries is already marketing tissue culture-derived shikonin, a red-colored naphthoquinone compound used as a dye and an astringent (21). Valuable pharmaceutical intermediates and drugs could in the future be produced in 1000-liter (and larger) vats of plant cells, a situation analogous to the current industrial production of antibiotics and other chemicals by continuous fermentation of microbial cultures (23, 52, 65).
Breeding and Cultivation of Plant Sources of Natural Products
Attention is often paid to the selection, breeding, and cultivation of plants from which natural products are obtained. Research in these areas frequently leads to vast improvements in yield of important plant-derived compounds. Tissue culture methodologies can be particularly useful in achieving rapid multiplication of the highest yielding individual plants. The potential success of the cultivation and improvement of commercially important plants may be illustrated by one example of how tissue culture techniques have been applied to this area. Elite genotypes of pyrethrum plants, whose flowers contain large amounts of pyrethrins, have been selected and propagated clonally by means of tissue culture techniques. Eleven million of these plants are now being grown in Ecuador. This approach potentially could be applied to other plants producing high-value metabolites.
Conclusions
Plant natural products have long been and will continue to be important sources and models of spices, flavors and fragrances, vegetable oils, soaps, natural rubber, gums, resins, drugs, insecticides, and other industrial, medicinal, and agricultural raw materials. For example, it is important to recall that the Industrial Revolution was made possible in part by the discovery and commercial development of the rubber tree, Hevea brasiliensis. However, since most plant species have never been described much less surveyed for chemical or biologically active constituents, it is reasonable to expect that new sources of valuable materials remain to be discovered. However, if the current trend of destruction of tropical forests continues at its present rate. phytochemists and other plant scientists may have only a few decades remaining in which to investigate much of the plant kingdom for useful chemicals.
Advances in chromatographic and spectroscopic techniques now permit the isolation and structural analysis of potent biologically active plant constituents that are present in quantities too minute to have been previously characterized. In addition. advances in plant cell and tissue culture and genetic manipulation could provide new means for economic production of rare plants and the chemicals that they produce. These new chemical and biological technologies will serve to enhance the continued usefulness of higher plants as renewable resources of chemicals.
M.F. Balandrin is Senior Natural Products Chemist, J.A. Klocke is Senior Entomologist-Natural Products Chemist, E.S. Wurtele is Senior Cell Biologist, and Wm. H. Bollinger is Vice President for Corporate Development at NPI (Native Plants, Inc.), University Research Park 417 Wakara Way, Salt Lake City, Utah 84108.
References and Notes
1. V.E. Tyler, L.R. Brady, J.E. Robbers, Pharmacognosy (Lea and Febiger, Philadelphia, ed. 8, 1981).
8. J. Meinwald, G.D. Prestwich, K. Nakanishi, I. Kubo, Science 199, 1167 (1978).
10. M. Jacobson, Econ. Bot. 36, 346 (1982).
11. J.B. Harborne, Introduction to Ecological Biochemistry (Academic Press, New York, ed. 2, 1982).
16. _____ and R.W. Morris, Am. J. Pharm. 148, 46 (1976).
19. J.D. Phillipson, Pharm. J. 222, 310 (1979); N. R. Farnsworth, Econ. Bot. 38, 4 (1984).
21. M.E. Crutin, Bio/Technology 1, 649 (1983).
22. K. Nakanishi, J. Nat. Prod. 45, 15 (1982).
23. Y. Aharonowitz and G. Cohen, Sci. Am. 245, 140 (March 1981).
25. P. Gund, J.D. Andose, J.B. Rhodes, G.M. Smith, Science 208, 1425 (1980).
26. D.A Hussar, Am. Pharm. NS24 (No. 3), 31 (1984).
29. L.W. Levy, Environ. Exp. Bot. 21, 389 (1981).
33. H. Gysin, Chimia 8, 205 (1954); E. Steadman, Biochem, J. 20, 719 (1926).
35. J.D. Cohen and W.J. Meudt, Plant Physiol. 72, 691 (1983).
37. M. Suffness and J. Douros, J. Nat. Prod. 45, 1 (1982); E. Frei III, Science 217, 600 (1982).
38. O.R. Gottleib and W.B. Mors, J. Agric. Food Chem. 28, 196 (1980).
40. V.E. Tyler, Econ. Bot. 33, 377 (1979).
45. J.A. Klocke, thesis, University of California, Berkeley (1982).
47. G.B. Staal, Annu. Rev. Entomol. 20, 417 (1975).
54. E. J. Staba; in Plant Tissue Culture 1982, A. Fujiwara, Ed. (Maruzen, Tokyo, Japan, 1982),
56. F. Sasse, U. Heckenberg, J. Berlin, Plant Physiol. 69, 400 (1982)
58. A. W. Altermann and E. Reinhard, unpublished data; see (53).
59. P. F. Heinstein, in Plant Tissue Culture 1982. A. Fujiwara, Ed. (Maruzen, Tokyo, Japan, 1982)
60. K. K. Kamo, W. Kimoto, A. F. Hsu, P. G. Mahlberg, D. D. Bills, Phytochemistry 21, 219(1982).
62. B. J. Nikolau, E. S. Wurtele, P. K. Stumpf, Plant Physiol. 75, 979 (1984).
{kind=link}
{kind=link}
{kind=link}