 Reproduced, with permission, from:
Neue, H. 1993. Methane emission from rice fields: Wetland rice fields may make a major contribution to global warming. BioScience 43 (7): 466-73.
Reproduced, with permission, from:
Neue, H. 1993. Methane emission from rice fields: Wetland rice fields may make a major contribution to global warming. BioScience 43 (7): 466-73.
Heinz-Ulrich Neue
Aside from being an important greenhouse gas, methane also affects the chemistry and oxidation capacity of the atmosphere, for example, by influencing concentrations of tropospheric ozone, hydroxyl radicals, and carbon monoxide. In the stratosphere, it is a sink for chlorine but a source for hydrogen and water vapor (ice crystals). The current burden of methane in the atmosphere is approximately 4700 Tg (1 Tg = 1 million tons; Wahlen et al. 1989), and the global annual emission is estimated to be 500 Tg with an apparent net flux of 40 Tg/yr (Cicerone and Oremland 1988).
The overall budget of atmospheric methane is fairly well established, but the strength of individual sources remains uncertain. Best estimates of sources and sinks from various reports are summarized in Table 1. Anthropogenic sources (340 Tg/yr) predominate over natural sources (160 Tg/yr), and 80% of the total methane emission is of modern biogenic origin. Only 20% is due to fossil carbon sources (Wahlen et al. 1989).
Wetland rice fields have recently been identified as a major source of atmospheric methane. Although the potential for methane release from rice fields has long been noted (Harrison and Aiyer 1913), the first comprehensive measurements of methane fluxes in rice fields were reported only in the early 1980s (Cicerone and Shetter 1981, Cicerone et al. 1983, Holzapfel-Pschorn et al. 1985, Seiler et al. 1984). As in a natural wetland, flooding a rice field cuts off the oxygen supply from the atmosphere to the soil, which results in anaerobic fermentation of soil organic matter. Methane is a major end product of anaerobic fermentation. It is released from submerged soils to the atmosphere by diffusion and ebullition and through roots and stems of rice plants. Recent global estimates of emission rates from wetland rice fields range from 20 to 100 Tg/yr (IPCC 1992), which corresponds to 6-29% of the total annual anthropogenic methane emission.
In this article, I discuss the importance of rice as a staple food, the different environments in which rice is grown, and the methane fluxes in rice fields. I describe the factors controlling those fluxes and options for mitigating methane release.
Conserving and enhancing wetland soils is essential to meet and sustain food production for future generations, as well as to conserve habitats for endangered plant and wildlife species. In Asia, wetlands have been used intensively for food production for thousands of years. In Africa and Latin America, wetlands are becoming a significant resource for food production. However, in Europe and North America, wetlands are being drained and turned into drylands with accelerating speed.
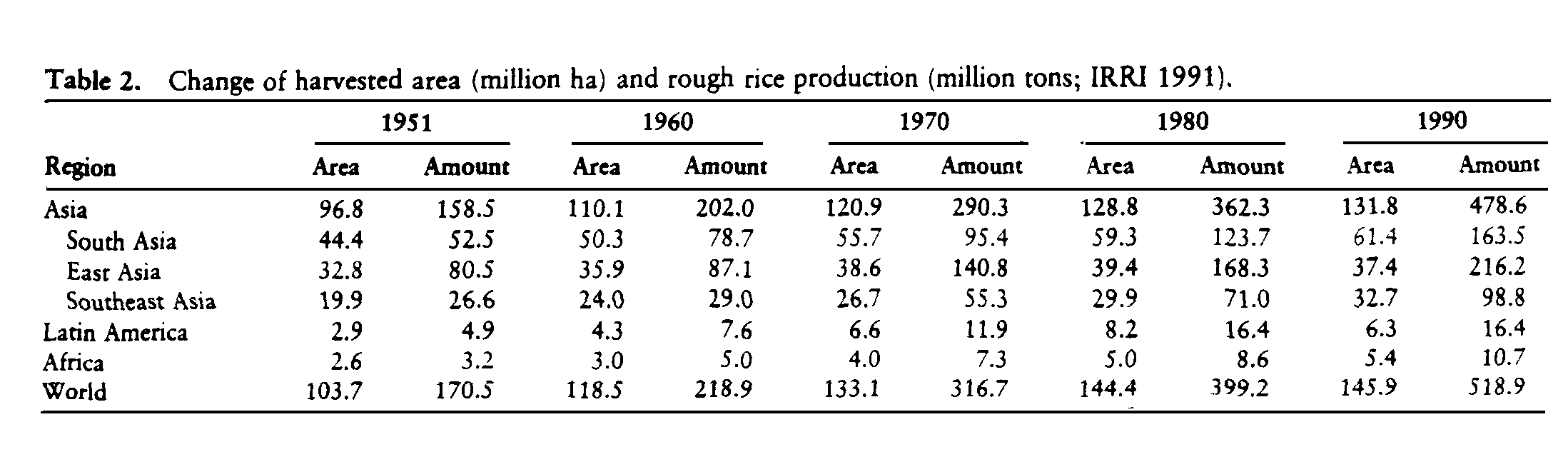
Rice is the world's most important wetland food crop, and pressure to grow more rice is accelerating. Rice is the only major grain crop that is grown almost exclusively as food. In 30 years, the earth may be home to 8 billion people, and the number expected to be rice consumers (the descendants of today's rice consumers) may equal today's total inhabitants of the earth. Feeding them will require a massive increase in global rice production. Making allowances for substitutions of other foods for rice in diets as incomes increase, the world's annual rice production must increase from 518 million tons in 1990 to 760 million tons in 2020 (IRRI 1989). That 47% increase would merely maintain current nutrition levels, which for hundreds of millions of people are already inadequate.
More than 90% of the world's rice is produced in Asia, 3.2% in Latin America (Brazil and Colombia account for 62% of that production), 2.1% in Africa (Egypt and Madagascar account for 48% of that production), and 2.5% in the rest of the world. Less than 5% of world rice production is traded on the international market. Rice provides between 35% and 59% of the calories consumed by 2.7 billion people in Asia. In Africa and Latin America, rice provides 8% of the food energy for almost 1 billion people. Approximately half the people in South Asia and subSaharan Africa receive inadequate calories for an active working life (IBRD 1986). Some 470 million of these malnourished people live in South Asia, and 150 million of them live in sub-Saharan Africa (IRRI 1989).
The world's harvested rice area increased during the past 40 years by 41% and rough (still in the hull) rice production by 304% (Table 2). Today, rice production in Asia is twice that of 25 years ago, whereas the land area planted to rice has increased only 17%. Average yields have increased 72%, which is only slightly more than the 67% growth in population (IRRI 1991).
The harvested rice area increased mainly because double and triple rice cropping was made possible by the development of short-duration, photoperiod-insensitive rice cultivars and by expanded irrigation. The great production gains have been in the irrigated rice and favorable rain-fed rice areas (Table 3), where modern high-yielding rice cultivars coupled with improved cultivation technologies are able to express their yield potential. In East Asia, almost all rice is grown on irrigated land; 40% of the rice-growing land in Southeast Asia is irrigated, but only 30% is irrigated in South Asia (Table 4).
There is a need to diversify agricultural systems on wetland soils to improve daily diet and to meet socioeconomic and environmental necessities for sustainable production systems in different regions. Various rice-based cropping and farming systems have evolved, and production of other staples (vegetables, plantation crops, livestock, poultry, and fish) are expanding into wetlands.
Wetlands are transitional between terrestrial and aquatic systems and generally include swamps, marshes, bogs, and similar areas. Wetlands occur where soils are naturally or artificially inundated or saturated by water due to high groundwater or surface water during part or all of the year. Wetlands are common in river deltas, estuaries, floodplains, and tidal areas, and they are widespread in river beds, depressions, footslopes, and terraces of undulating landscapes.
Ricelands can be classified as irrigated, rain-fed, deepwater, or upland. In irrigated rice fields, the floodwater is fully controlled and kept shallow. In rain-fed rice fields, precipitation controls flooding of soils. At times in the growing season, soils of rain-fed rice fields may dry up or be flooded up to 50 cm. In deepwater rice fields, floodwater rises to more than 50 cm during the growing season, and it may reach several meters. Upland rice fields are neither flooded nor does the topsoil become water saturated at any significant period of time. Most rice is grown on wetlands; only 13% is cultivated on uplands as is wheat or malze.
Wetland rice fields have at least one wet growing season, but they may be without surface water, moist, or dry in other seasons. These fields may therefore alternately support wetland and upland crops and, in fact, normally do. The boundary between wetland and upland is often gradual and may fluctuate from year to year depending on variations in precipitation. If water (drainage and irrigation) can be fully controlled, it is within a farmer's discretion to establish wetland or upland crops. But in most wetland rice areas, drainage is insufficient to prevent soil submergence during the rainy season.
Wetland rice soils have free water at the surface for at least the major part of the crop growing season. Water may be retained by leveling and building levees or dikes. Flooding drastically reduces the diffusion of atmospheric oxygen into the soil, and facultative and anaerobic microorganisms sequentially reduce soil substrates. The redox potential is a quantitative indicator measuring the tendencies of different oxidations and reductions (net flow of electrons) to occur. Redox potential, measured as electric potential in volts, characterizes the processes that bring about a given chemical and biochemical milieu in a soil. The higher the value of the redox potential, the greater the presence of strong oxidizing agents in a soil.
The reduction of soils upon flooding proceeds stepwise in a thermodynamic sequence. Free oxygen is reduced first, followed, in order, by nitrate, manganic manganese compounds, ferric iron compounds, sulfate, and, last, carbon dioxide. Consequently, the redox potential falls sharply and the partial pressure of carbon dioxide increases considerably. As a result, the pH of acid as well as of alkaline soils stabilizes at about neutral, and availability of most nutrients increases.
The anaerobic fermentation of organic matter produces and accumulates an array of substances, many of them transitory and not found in well aerated soils. These substances include various gases, hydrocarbons, alcohols, carbonyls, volatile and nonvolatile fatty acids, phenolic acids, and volatile sulfur compounds. The degree of humification (polymerization of organic compounds to humic substances) is low, but the decomposition of organic matter can be as rapid as in upland soils. The magnitude of reduction is determined by the amount of easily degradable organic matter, the rate of decomposition, the formation of toxins to microorganisms, and the amounts and kinds of reducible nitrates, manganese and iron oxides, sulfates, and organic compounds. The most important redox buffer system in wetland soils is comprised of iron and organic compounds. Carbon dioxide and methane are the end products of organic matter decomposition in wetland rice soils.
A flooded rice soil has a reduced topsoil with an oxidized floodwater-soil interface. The depth of the interface is usually between 2 and 20 mm, depending on the quantity of oxygen dissolved in the floodwater, the reducing capacity of the soil (carbon content), the water percolation, and the activity of the soil and water fauna (Neue 1991). Major processes in the interface include aerobic decomposition of organic matter, photodependent biological nitrogen fixation by algae and photosynthetic bacteria, nitrification by ammonium and nitrite oxidizers, and methane oxidation (Watanabe and Furasaka 1980). In the reduced topsoil, anaerobic decomposition of organic matter, heterotrophic biological nitrogen fixation, denitrification, manganese reduction, organic acid production, iron reduction, sulfate reduction, hydrogen production, and methanogenesis occur.
A soil layer called a plow pan may develop between the topsoil and the subsoil due to cultivation. It exhibits lower permeability and higher bulk density and mechanical strength than other soil layers. Plow pans reduce detrimental water percolation and nutrient leaching. The subsoil below the plow pan is aerobic when flooding occurs by surface water (epiaquic moisture regime) and anaerobic in soils where groundwater rises to the surface.
Flooding reduces the physical variability of soils, increases its fertility, and confers stability to crop systems that, like rice, can tolerate standing water. Flooding a soil provides an ideal growth medium by supplying abundant water that may carry significant amounts of suspended solids and nutrients, buffering soil pH near neutral, enhancing nitrogen fixation and carbon supply, and increasing diffusion rates, mass flow, and availability of nutrients. Standing water stabilzes the soil moisture regime, moderates soil temperature, and prevents soil erosion. Flooding a soil depresses soil-borne plant diseases and weed growth.
Methane is produced as the terminal step of the anaerobic breakdown of organic matter in wetland rice soils. Methane is exclusively produced by methanogenic bacteria that can metabolize only in the strict absence of free oxygen and at redox potentials of less than -150 mV (Wang et al. in press). Most methanogens are neutrophilic, with an optimal pH of 6-8. Methanogens rely on a plethora of other microorganisms to provide them with the few substrates they can catabolize: hydrogen, carbon dioxide, formate, acetate, methanol, methylamines, and methysulfides (Conrad 1989, Garcia 1990). In wetland rice soils, methane is largely produced by transmethylation of acetic acid and, to some extent, by the reduction of carbon dioxide (Takai 1970).
The formation of methane is preceded by the production of volatile acids. On flooding, short-term evolution of hydrogen immediately follows the disappearance of oxygen, carbon dioxide increases, and, with decreasing carbon dioxide, methane formation increases (Neue and Scharpenseel 1984, Takei et al. 1956). The delay of methane production depends on the pattern of soil reduction, pH, substrate availability, and temperature. In tropical flooded rice soils, where soil temperatures are 25-30deg.C, methane production in alkaline and calcareous soils may start hours after flooding, in neutral soils it is delayed two to three weeks, and in acid soils methane may only be formed five or more weeks after flooding. Methane production is negatively correlated with e soil-redox potential and positively correlated with soil temperature, soil carbon content, and rice growth (Neue and Roger in press).
The rate and pattern of organic matter addition and decomposition determine the rate and pattern of methane formation. Methane production generally increases during the cropping season, although the population density of methanogens remains fairly stable (Schutz et al. 1989b). Easily degradable crop residues, fallow weeds, and soil organic matter are the major source for initial methane production. At later growth stages of rice, root exudates, decaying roots, and aquatic biomass seem to be more important. Methane production is enhanced in the rooted soil zones (Sass et al. 1991).
Inhibition of methane formation
Decomposition of organic matter and methane production is retarded in wetland soils with acidic or allic reaction, low and imbalanced nutrient supply, high kaolinitic clay content, and/or high bulk density (Neue and Scharpenseel 1987, Neue et al. 1990, Neue and Roger in press). Sodium chloride inhibits methanogenesis when added in high (approximately 0.18 M) concentrations (Patel and Roth 1977). Addition of sea water inhibits methane formation at lower salt concentrations because of its sulfate content (Koyama et al. 1970). Competition of sulfate-reducing bacteria for hydrogen and hydrogen sulfide toxicity are the likely mechanism. However, methanogenesis and sulfate reduction are nor mutually exclusive when methane is produced from methanol and methylated amines for which sulfate reducers show little affinity (Oremland et al. 1982).
Some halogenated organic compounds--chloroform, DDT, acetylene, and nitrapyrin (an inhibitor of nitrification)--completely inhibit the production of methane (Bauchop 1967, McBride and Wolfe 1971, Raimbault 1975, Salvas and Taylor 1980, Smith and Mah 1981). Activity in each case is believed to depend on the presence of a trichloromethyl group that inhibits the action of methyl coenzyme M reductase. Resistance to bromoethanesulfonate does not confer resistance to a number of chlorinated inhibitors (Smith and Mah 1981). Ammonium toxicity using Methanobacteriun formicium has been reported by Hobson and Shaw (1976).
Methane-oxidizing bacteria (methanotrophs) are abundant in the oxidized floodwater-soil interface and in the rice rhizosphere. They sequentially oxidize methane to carbon dioxide via methanol, formaldehyde, and formate. Oxygen is essential for the growth of methanotrophs, but the required partial pressure may be low (Cicerone and Oremland 1988). Methane oxidation greatly limits diffusion of methane to the atmosphere. Up to 60% of the methane produced during a rice growing season may be oxidized before it reaches the atmosphere (Holzapfel-Pschorn et al. 1986, Sass et al. 1991). Ammonium ion inhibited methane oxidation in studies with pure cultures of methanotrophs (Hyman and Wood 1983, Whittenbury et al. 1970). Field experiments have revealed no significant effect of ammonium ions, probably because of their immediate uptake by rice plants.
Rice plants supply atmospheric oxygen to the roots for respiration via a special vascular system, the aerenchyma. The aerenchyma has its own openings at the leaf sheath (Nouchi et al. 1991), and the gas supply to and from the roots is independent of transpiration and stomatal gas exchange. Oxygen diffusion from rice roots constitutes an important part of the roots' oxidizing power, aside from enzymatic hydrogen peroxide production. Because of the abundance of methane-oxidizing bacteria present in the rhizosphere, the rhizophere's potential for methane oxidation is high.
De Bont et al. (1978) counted ten times more methane-oxidizing bacteria in the rhizosphere than in the bulk anaerobic soil and one-third more than in the oxidized soil-water interface. They found significant increases in methane emission by the rice cultivar IR36 when methane oxidation was suppressed with acetylene at the soilwater interface. However, acetylene had only a small effect on emission rates when applied to the rhizosphere. De Bont and his colleagues concluded that the use of oxygen by reduced substances and microbes other than methanotrophs at the region of the root-soil interface exceeds the supply of oxygen by the root. Consequently, the aerobic zone surrounding the root of IR36 is too thin to oxidize the diffusing methane, or the rhizosphere is for the most part anaerobic. Nevertheless, variability in root-oxidizing power of rice cultivars is high, and the impact of roots on methane oxidation merits further study.
Methane is released from anaerobic wetland soils to the atmosphere through diffusion of dissolved methane, ebullition of gas bubbles, and via plants that, like rice, develop aerenchyma tissue. Large portions of methane formed in an anaerobic soil may remain trapped in the flooded soil. Entrapped methane may be oxidized to carbon dioxide when the floodwater is drained during the rice growing season or when the soil dries at the end of or after the rice growing season. But large amounts of entrapped methane may escape to the atmosphere immediately after the floodwater recedes (Denier van der Gon et al. 1992).
The low solubility of methane in water limits its diffusive transport in the flooded soil, and most methane is oxidized to carbon dioxide via methanol, formaldehyde, and formate as it passes the aerobic soil-water interface. The release of methane by diffusion through the wet soil column is negligible in clayey soil, but it may become significant in sandy soils in which bigger pores between soil particles prevail. Most rice soils have high clay contents. Soil fauna, especially aquatic earthworms (Tubificidae), increase emission through diffusion and ebullition when they dig into the topsoil. At the same time, oxidation of methane is enhanced. In deepwater rice fields, diffusing methane may only be oxidized in the upper water column, because the soil-water interface and the lower water column may be anaerobic.
Usually, rice fields are prepared by flooding two to four weeks before rice is transplanted. If bare mud is flooded, most methane is trapped in the soil, and as long as the soil is not heavily amended with organic matter and remains undisturbed, only small amounts of methane are released through ebullition. Under actual field conditions, large portions of the soiltrapped methane escape to the atmosphere through ebullition caused by such culture practices as wet tillage, puddling, harrowing, transplanting, fertilization, weeding, pest management, and harvest. In fields planted to wetland rice and not disturbed by culture practices during the growing season, up to 70% of the methane released to the atmosphere is emitted through the aerenchyma of the rice plant. During the reproductive phase, more than 90% of the emission is attributed to the rice plant (Cicerone and Shetter 1981, Schutz et al. 1989b).
Field measurements of methane emission from wetland rice fields have been conducted in the United States ( Cicerone and Shetter 1981, Lindau et al. 1991, Sassetal. 1991),Spain (Seiler et al. 1984), Italy (Holzapfel-Pschorn et al. 1986, Schutz et al. 1989a),China (Khalil et al. 1991, Schutz et al. 1990), Japan (Yagi and Minami 1990), and the Philippines (IRRI 1992). As with methane production, methane emission rates are highly variable. A distinct diel and seasonal pattern is observed. Emission rates mediated by rice plants are highest in the early afternoon and lowest late at night, following the diel pattern of air and topsoil temperature and pH as well as oxygen concentrations in the floodwater and soil-water interface. The control mechanisms have still to be elucidated and may be more complex because diel amplitudes of the above factors vary independently over the growing season. The basic level and the diel amplitudes of methane emission increase with addition of organic matter.
Two or three maxima of plant mediated emission rates are generally observed in irrigated rice fields during the cropping season. Methane emission increases during the vegetative phase, sometimes with an early intermediate peak; it decreases after panicle initiation and increases again after flowering, before declining at the end of the season. For irrigated rice, methane emissions calculated as a function of rice produced are higher in the dry season than in the wet season. Various climatic, soil, water, and culture factors can modify this general pattern.
Organic amendments increase methane production and emission. Composted materials enhance methane fluxes less than do green manure or rice straw. Composting depletes easily degradable carbon and transforms organic substrates into more stable humus. Application of chemical fertilizer improves plant growth and therefore increases methane emission and probably its production. Fertilizer containing sulfate may enhance growth and reduce methane production even if the emission apparently increases, because of better plant growth. Intermittent drying of flooded rice fields reduces total methane production and emission (Kimura 1992, Sass et al. 1992).
Soil disturbances during culture increase ebullition to methane. Broadcasting germinated seeds on wet cultivated fields may emit less methane than transplanting rice. The growth duration of direct wet-seeded rice and consequently its flooding period is shorter. Flooding periods are even shorter when rice is directly seeded into dry soils, as it is in Australia and the United States. Release of entrapped methane through soil disturbance is less frequent in direct-seeded rice because there are more plants per unit area, weed growth is suppressed, and hand or mechanical weeding becomes impractical.
Global estimates of methane emission rates from rice fields
Irrigated rice fields are the major source of methane from rice fields. Although irrigated rice comprises only 50% of the harvested rice area, it produces 70% of the rice harvested. Methane production is favored by the assured water supply and control, intensive soil preparation and fertilization, and resultant improved growth of the rice.
Methane emissions are much lower and more variable in rain-fed rice because of drought periods during the growing season and poorer growth of rice. In deepwater rice, methane production may be high, but related emission rates may be low because of reduced emission pathways. Upland rice is not a source of methane emission because it is not flooded for any significant period of time.
Because of the limited number and locations of comprehensive seasonal flux measurements, global extrapolations of emission rates are highly uncertain and tentative. Recent global methane budgets from rice fields range from 20 to 100 Tg/yr. The global flux is usually estimated from the harvested wetland rice area at different regions, seasonal mean emission rates, and length of the emission period. Major problems arise in deriving the seasonal mean emission rates and the length of emission periods for regions. At present, there are insufficient data and knowledge to account for variations of climate, soil properties, duration and pattern of flooding, organic amendments, fertilization, other culture practices, and rice growth.
By making different assumptions about effects of soil types, temperature, recycling of crop residues, rice growth, and length of rice growing seasons, and accordingly adjusting a basic emission rate of 0.5 g m2 day-1 for wetland rice areas in each country, the source strength of rice fields in Asia was estimated to range between 40 and 60 Tg/yr (Bachelet and Neue in press). Comprising 51% of the global harvested rice area, rice fields in China and India emit methane at a rate of only 26-37 Tg/yr.
Current global estimates of methane fluxes from rice fields do not account for varying floodwater regimes, especially in rain-fed rice areas where drought spells may drastically lower emission rates. Nor do they account for organic amendments, ebullition induced by culture, and release of entrapped methane during initial drying phases, which should increase emission rates. Unless factors and processes of methane emission are well understood and simulated mechanistically, estimates of global methane emission from rice fields and responses to climate change or changes in cultivation practices will remain controversial.
Mitigation options
The most effective mitigation option would be to prevent submergence of rice fields and to cultivate upland rice or other upland crops. However, there are two major problems with such a scheme. First, wetland rice is usually grown because fields are flooded naturally during the rainy season, not vice versa. Drainage of rice fields is often impossible during rainy seasons. Impounding water in rice fields is also an important factor in soil erosion control at areas of high precipitation and groundwater regimes in areas of monsoon Asia that have distinct dry and wet seasons. Wetland rice fields are excellent substitutes for tropical rainforests with regard to water and erosion control and sustained soil fertility. Second, upland rice would not be a good replacement for wetland rice because its production potential at the same levels of inputs is much lower. Mitigation options must achieve both reduced methane emission and increased sustainable production of rice.
Water management. Increasing water percolation would add oxygen-rich water to the reduced soil layer and decrease methane production. However, such mitigation practice is only feasible where complete control of water supply and drainage is possible. In drought-prone rain-fed areas, water is too valuable to be drained. Higher water percolation requires more water and may cause detrimental leaching of nutrients. Significant quantities of methane may also be leached and subsequently released to the atmosphere elsewhere (Kimura 1992). Temporarily aerating the soil by stopping irrigation enhances methane oxidation and decreases methane formation as well as total emission. But intermittent aeration may increase gaseous losses of nitrogen as nitrous oxide (nitrification-dinitrification). Drying of rice terraces at hillsides can cause severe cracking and collapse of the terrace construction.
Water stress at any growth stage reduces rice yield. A soil moisture content of -50 kPa (slightly above field capacity) may reduce rice grain yield by 20-25% compared to continually flooded treatments (De Datta 1981). The rice plant is most sensitive to water stress during the reproductive stage, when lack of water causes a high percentage of sterility (Yoshida 1981). Water deficit during the vegetative stage reduces plant height, tiller number, and leaf area, and it sharply reduces yields if plants do not recover before flowering.
The duration of a moisture stress is more important than the growth stage at which the stress occurs. Intermittent drying or keeping soils only saturated during the growing season considerably lowers rice yields in most tropical rice fields (Borell et al. 1991). But in subtropical China, Japan, and Korea, intermittent drying periods as well as percolation rates of up to 35 mm/day are associated with maximal rice yields, probably because organic and inorganic toxins accumulate due to low soil temperature early in the season. Short aeration periods at the end of the tillering stage and just before heading improve wetland rice yields (Wang Zhaoqian 1986) only if followed by flooding.
Water management is a promising candidate to mitigate methane emission from rice fields. But sound and feasible management practices that reduces methane emissions, without enhancing nitrogen losses and increasing rice yields, must be developed.
Rice cultivars. The aerenchyma of rice plants mediate the transport of air (oxygen) to the roots and methane from the anaerobic soil to the atmosphere. The flux of gases in the aerenchyma depends on concentration gradients and diffusion coefficients of roots and internal structure, including openings of the aerenchyma. The number of tillers per area, the root mass, the rooting pattern, and metabolic activity also influence the gas fluxes. Oxygen diffusion and exudation of oxygen radicals combined with abundant methane-oxidizing bacteria result in oxidization of methane in the rhizosphere, whereas organic root exudates and root litter are a source of methane formation.
The wide variation of these traits and related emission rates among cultivars opens the possibility for breeding rice cultivars with low methane emission potential. The inheritance of underlying traits and relationships to yield potential have yet to be elucidated.
Fertilization and other cultivation practices. Quality as well as quantity of added organic materials influence methane formation. Amendments with fresh organic matter, like rice straw and green manures, highly increase methane production and emission. Application of composted material, which has a higher degree of humification, only slightly increases methane formation and fluxes (Yagi and Minami 1990). Sound technologies must be evaluated with consideration of maintaining and increasing soil fertility as well as mitigating methane emission. To reduce methane emission from wetland rice fields, it may be necessary to minimize rather than maximize organic amendments. But green manures and recycling of crop residues are sometimes the only soil conditioner and nutrient source for many resource-poor farmers. In general, use of organic amendments declines as chemical fertilizers, especially nitrogen fertilizer, become available and responsive rice cultivars are grown.
Urea accounts for approximated 80% of the nitrogen applied to rice in Asia, and ammonium sulfate accounts for approximately 6% (De Datta 1981). Most farmers apply nitrogen fertilizer in two or three splits. The first split is applied during final land preparation or shortly after planting, and the remainder is broadcast at later growth stages, especially at panicle initiation. To minimize volatilization losses of nitrogen fertilizer, it is recommended that fertilizer granules be placed 10 cm deep in the soil during the final land preparation. In general, potassium and phosphorus fertilizer are basal-applied during final land preparation. Potassium chloride is the principal fertilizer source of potassium, and superphosphate is the primary source of phosphorus fertilizer. On acid rice soils, phosphate rock may be applied.
The direct impact of chemical fertilizer applications on methane emission is not clear. Because most methane is emitted through the rice plant, improved rice growth (more tiller and roots) in response to fertilizer application increases emission. But source and mode of application may also have direct effects (Schutz et al. 1989a). Sulfate-containing fertilizer reduces methane emission. Sulfate-reducing bacteria compete with methanogens for the limited hydrogen, but the amount of sulfate normally added as fertilizer seems to be insufficient to have significant effects.
Nitrification inhibitors, such as nitrapyrin and acetylene, incorporated into the soil also limit methane production without reducing much methane oxidation. Slow release of acetylene from urea-encapsulated calcium carbide greatly reduced methane and nitrous oxide emissions from rice in greenhouse experiments and increased rice yield (Bronson and Moiser 1991).
Impacts of various other culture practices (land preparation, seeding and transplanting, pest control, and harvest) on methane emission have not yet been studied in detail. A few observations at the International Rice Research Institute (IRRI) reveal that soil disturbances caused by current culture practices release large amounts of soil-entrapped methane. The increased adoption of direct seeding (wet and dry seeding) instead of transplanting are likely to reduce methane emission. In direct-seeded rice, flooding periods are shorter and cultural disturbance of reduced soils is minimized.
Rice-based cropping systems. Crop diversification is a feasible option to reduce total methane emission and increase production as well as income (Neue et al. 1991). In rice growing areas with vear-round irrigation, production can be increased by growing two to three rice crops each year or by sequentially cropping an upland crop before or after one or two crops of rice. In rain-fed wetland areas, where residual moisture is sufficient, the wet season rice crop may be followed by an upland crop. Most rain-fed areas remain fallow in rhe dry season.
The most common upland crops grown in rotation before rice are early maturing corn, mung bean, cowpea, and potato. Crops grown after rice are wheat, mustard, mung bean, cowpea, soybean, corn, chickpea, sweet potato, lentil, and black grass (Morris and et al. 1986). Crop intensification through crop sequencing has become possible because early maturing varieties allow farmers to gain as much as 45 days in a growing season for additional crop production. Direct-seeded rice can even be harvested 15-30 days earlier than transplants of the same variety. Residual soil moisture after wetland rice culture often can produce a short-duration, well-rooted upland crop, if care is taken to reduce water loss during land preparation and the early vegetative stage of upland crops (i.e., minimal or no tillage; Gomez and Zandstra 1982).
In wetland rice-based cropping systems, soils are cultivated in distinct cycles of anaerobic and aerobic phases. This cycle implies transformation from a puddled to a granular soil structure and radical alterations in nutrient kinetics associated with changes in the redox status of the soil. Establishment of upland crops following wetland rice is a major problem in cropping intensification. Excess water may delay land preparation, and puddled soils, especially clayey soils, become compact and hard on drying. Upland crops also stand the risk of flooding when heavy late rainfalls occur.
Conclusions
The complex interactions among methane formation, methane oxidation, rice growth and cultivation, and methane emission require an immediate, integrated, and interdisciplinary research approach, including application of socioeconomics and participation of farmers, to achieve the knowledge needed to design feasible and effective mitigation technologies. With current cultivation technologies, methane emission from rice fields is expected to increase, as rice production is increased by 50 to 100% within the next three decades. By using a combination of feasible mitigation technologies, however, there is great potential to stabilize or even reduce methane emission from rice fields while increasing rice production, without dramatically changing culture practices.
With financial support from the US Environmental Protection Agency, IRRI has started baseline research on methane fluxes in rice fields in collaboration with the Fraunhofer Institute for Atmospheric Environmental Research, in Garmisch-Partenkirchen, Germany, and the Wetland Biogeochemistry Institute of Louisiana State University in Baton Rouge. To support national rice research systems in developing their own capacity to respond to the issue of methane emission from rice fields and link their expertise with that of advanced institutions, IRRl is also coordinating an interregional research program on methane emission from rice fields funded by the Global Environmental Facility of the United Nations Development Programme. This program comprises collaborative methane research on irrigated, rain-fed, and deepwater rice in China, India, Indonesia, Philippines, and Thailand.
References cited
Bachelet, D., and H.-U. Neue. In press. Methane emissions from wetland rice areas of Asia. Chemosphere.
Bauchop, T. 1967. Inhibition of rumen methanogensis by methane analogues. J. Bacteriol. 94: 171.
Borell, A. K., S. Fukai, and A. L. Garside. 1991. Irrigation methods for rice in tropical Australia. Int. Rice Res. News. l6(3): 28.
Bouwman,A.F.,ed. 1989. Soils and the Greenhouse Effect. John Wiley & Sons. New York .
Bronson. K. F., and A. R. Mosier.1991. Effect of encapsulated calcium carbide on dinitrogen, nitrous oxide, methane and carbon monoxide emissions in flooded rice. Biol. Fertil. Soils 7: 116-120.
Cicerone, R. J., and R. S. Oremland. 1988. Biogeochemical aspects of atmospheric methane. Global Biochem. Cycles 2: 299-327.
Cicerone, R. J., and J. D. Shetter.1981. Sources of atmospheric methane: measurements in rice paddies and a discussion. J. Geophys. Res. 86: 7203-7209.
Cicerone R. J., J. D. Shetter, and C. C. Delwiche. 1983. Seasonal variation of methane flux from a Californian rice paddy. J. Geophys. Res. 88: 11022-11024.
Conrad, R. 1989. Control of methane production in terrestrial ecosystems. Pages 39-58 in M. O. Andrae and D. S. Schimmel, eds. Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere. John Wiley & Sons, New York.
de Bont, J. A. M., K. K. Lee, and D. F. Bouldin. 1978. Bacterial oxidation of methane in rice paddy. Ecol. Bull. 26: 91-96.
De Datta, S.K. 1981. Principles and Practices of Rice Production. John Wiley & Sons, New York.
Denier van der Gon, H. A. C., H. U. Neue, R. S. Lantin, R. Wassmann, M. C. R. Alberto, J. B. Aduna, and M. J. P. Tan. 1992. Controlling factors of methane emission from ricefields. Pages 81-92 in N. H. Batjes and E. M. Bridges, eds. World Inventory of Soil Emission Potentials. World Inventory of Soil Emission Potentials Report 2, International Soil Reference and Information Center, Wageningen, The Netherlands.
Dickinson, R. E., and R. J. Cicerone. 1986 Future global warming from atmospheric trace gases . Nature 319: 109-115.
Garcia, J. L. 1990. Taxonomy and ecologv of methanogens. FEMS Microbiol. Rev. 87: 297-308.
Gomez, A. A., and H. G. Zandstra. 1982. Research strategy for rice-based cropping systems. Pages 381-394 in Rice Research Strategies for the Future. International Rice Research Institute, Los Baños, Philippines.
Harrison, W. H., and P. A. S. Aiyer. 1913. The gases of swamp rice soil.1. Their composition and relationship to the crop. Memoires Department of Agriculture India Chemistry Series 5(3): 65-104.
Hobson, P. N., and B. G. Shaw. 1976. Inhibition of methane production by Methanobacterium formicium. Water Res. 10: 849.
Holzapfel-Pschorn, A., R. Conrad, and W. Seiler. 1985. Production, oxidation and emission of methane in rice paddies. FEMS Microbiol. Ecol. 31: 343-351.
______. 1986. Effects of vegetation on the emission of methane from submerged paddy soils. Plant & Soil 92: 223--233.
Hyman, M., and P. M. Wood. 1983. Methane oxidation by Nitrosomonas europaea. Biochem.J. 212: 31-37.
Intergovernmental Panel on Climate Change (IPCC).1992. Climate Change: TheSupplementarv Report to the IPCC Scientific Assessment. Cambridge Universitv Press, New York.
International Bank for Reconstruction and Development/The World Bank (IBRD). 1986. Povert- and Hunger: A World Bank Policy Study. Washington, DC.
Inrernational Rice Research Institute (IRRI). 1989. IRRI Towards 2000 and Beyond. IRRI, Manila, Philippines.
_____. 1991. World Rice Statistics l990. IRRI, Manila, Philippines.
_____. 1992. Program Report for 1991. IRRI, Manila, Philippines.
Khalil, M. A. K., R. A. Rasmussen, M. X. Wang, and L. Ren.1991. Methane emission from rice fields in China. Environ. Sci. Technol. 25: 979-981.
Khalil, M. A. K., and M. Shearer, eds. In press. Global Atmospheric Methane. Springer-Verlag, New York.
Kimura, M. 1992. Methane emission from paddy soils in Japan and Thailand. Pages 43-79 in N. H. Batjes and E. M. Bridges, eds. World Inventory of Soil Emission Potentials. WISE Report 2, ISRIC, Wageningen.
Koyama, T., M. Hishida, and T. Tomino. 1970. Influence of sea salts on the soil methabolism. 11. On the gaseous metabolism. Soil Sci. PlantNutr. 16: 81-86.
Lindau, C. W., P. K. Bollich, R. D. Delaune, W. H. Patrick Jr., and V. J. Law.1991. Effect of urea fertilizer and environmental factors on methane emissions from a Louisiana, USA rice field. Plant & Soil 136: 195-203.
McBride, B. C., and R. S. Wolfe. 1971. Inhibition of methanogenesis by DDT. Nature 234: 551.
Morris, R. A., A. Patanothai, A. Syarifuddin, and V. R. Carangal. 1986. Associated crops in rice-based cropping systems. Pages 59-76 in Progress in Rainfed Lowland Rice. International Rice Research Institute, Manila, Philippines.
Neue, H.-U. 1991. Holistic view of chemistry of flooded soil. Pages 3-32 in Soil Management for Sustainable Rice Production in the Tropics. International Board for Soil Research and Management Monograph No.2, Bangkok, Thailand.
Neue, H.-U., P. Becker-Heidmann. and H. W. Scharpenseel. 1990. Organic matter dynamics, soil properties and cultural practices in
rice lands and their relationship to methane production. Pages 457-466 in A. F. Bouwman, ed. Soils and the Greenhouse Effect. John Wiley & Sons, New York.
Neue, H.-U., and P. A. Roger. In press. Rice agriculture: factors controlling emissions. In M. A. K. Khalil and M. Shearer, eds. Global Atmospheric Methane. Springer-Veriag, New York.
Neue, H.-U., C. Quijano, and H. W. Scharpenseel. 1991. Conservation and enhancement of wetland soils. Pages 279-303 in Evaluation for Sustainable Land Management in the Developing World. International Board for Soil Research and Management Proceedings No. 12(2), Bangkok, Thailand.
Neue, H.-U., and H. W. Scharpenseel. 1984. Gaseous products of the decomposition of organic matter in submerged soils. Pages 311-328 in Organic Matter and Rice. International Rice Research Institute, Manila, Philippines.
_____.1987. Decomposition pattern of 14C-labelled rice straw in aerobic and submerged rice soils of the Philippines. Sci. Total Environ. 62: 431-434.
Nouchi, I., S. Mariko, and K. Aoki. 1991. Mechanisms of methane transport from the rhizospere to the atmosphere through rice plants. Plant Physiol. 94: 59-66.
Oremland, R. S., L. M. Marsh, and S. Polcin. 1982. Methane production and simultaneous sulfate reduction in anoxic saltmarsh sediments. Nature 296: 143-145.
Patel, G. G., and L. A. Roth. 1977. Effect of sodium chloride on growth and methane production of methanogens. Can. J. Microbiol. 6:893.
Raimbult, M. 1975. Etude de l'influence inhibitrice de l'acetylene sur la formation biologique du methane dans un sol de riziere. Ann. Microbiol. (Inst. Pasteur) 126a: 217-258.
Ramanathan, V., R. J. Cicerone, H. B. Singh and J. T. Kiehl. 1985. Trace gas trends and their potential role in climate change. J. Geophys. Res. 90: 5547-5566.
Salvas, P. L., and B. F. Taylor. 1980. Blockage of methanogenesis in marine sediments by the nitrification inhibitor 2-chloro-6 (trichloromethyl) pyridine (Nitrapin or N-serve). Curr. Microbiol. 4: 305.
Sass. R. L., F. M. Fischer, P. A. Harcombe, and F. T. Turner. 1991. Methane production and emission in a Texas rice field. Global Biogeochem. Cycles 4: 47-68.
Sass, R. L., F. M. Fischer, Y. B. Wang, F. T. Turner, and M. F. Jund. 1992. Methane emission from rice fields: the effect of floodwater management. Global Biogeochem. Cycles 6: 249-262.
Schütz, H., A. Holzapfel-Pschorn, R. Conrad, H Rennenberg, and W. Seiler. l989a. A 3-year continuous record on the influence of daytime, season and fertilizer treatment on methane emission rates from an Italian rice paddy field. J. Geophys. Res. 94: 16405-16416.
Schütz, H., W. Seiler, and R. Conrad. 1989b. Processes involved in formation and emission of methane in rice paddies. Biogeochemistry 7: 33-53.
Schütz, H., W. Seiler, and H. Rennenberg. 1990. Soil and land use related sources and sinks of methane (CH4) in the context of the global methane budget. Pages 268-285 in A. F. Bouwman, ed. Soils and the Greenhouse Effect. John Wiley & Sons, New York.
Seiler, W., A. Holzapfel-Pschorn, R. Conrad, and D. Scharffe. 1984. Methane emission from rice paddies. J. Atmos. Chem. 1: 241-268.
Smith, M. R., and R. A. Mah. 1981. 2-Bromoethanesulfonate: a selective agent for isolating resistant Methanosarcina mutans. Curr. Microbiol. 6: 321.
Takai, Y. 1970. The mechanism of methane fermentation in flooded soils. Soil Sci. Plant Nutr. 16: 238.
Takai, Y., T. Koyama, and T. Kamura. 1956. Microbial metabolism in reduction process of paddy soil. Part 1. Soil Plant Food 2(2): 63-66.
Wahlen, M., N. Tanaka, R. Henry, B. Deck, J. Zeglen, J. S. Vogel,J. Southon, A. Shemesh, R. Fairbanks, and W. Broecker. l989. Carbon-14 in methane sources and in atmospheric methane: the contribution from fossil carbon. Science 245: 286-290.
Wang Zhaoqian. 1986. Rice based systems in subtropical China. Pages 195-206 in A. S.
{kind=link}
{kind=link}
{kind=link}
{kind=link}