 Reproduced, with permission, from:
Goward, S. N., C. J. Tucker, and D. G. Dye. 1985. North American vegetation patterns observed with the NOAA-7 Advanced Very High Resolution Radiometer. Vegetatio 64: 3-14.
Reproduced, with permission, from:
Goward, S. N., C. J. Tucker, and D. G. Dye. 1985. North American vegetation patterns observed with the NOAA-7 Advanced Very High Resolution Radiometer. Vegetatio 64: 3-14.
Samuel N. Goward1*, Compton J. Tucker2 & Dennis G. Dye1
1Department of Geography, University of Maryland, College Park, MD 20742 USA
2 Earth Resources Branch, NASA/Goddard Space Flight Center, Greenbelt, MD 20771 USA
Keywords: Continental-scale vegetation observation, Spectral vegetation index, NOAA-7 Advanced Very High Resolution Radiometer, Seasonality, Integrated growing season, Net primary productivity
Abstract Spectral vegetation index measurements derived from remotely sensed observations show great promise as a means to improve knowledge of land vegetation patterns. The daily, global observations acquired by the Advanced Very High Resolution Radiometer, a sensor on the current series of U.S. National Oceanic and Atmospheric Administration meteorological satellites, may be particularly well suited for global studies of vegetation. Preliminary results from analysis of North American observations, extending from April to November 1982, show that the vegetation index patterns observed correspond to the known seasonality of North American natural and cultivated vegetation. Integration of the observations over the growing season produced measurements that are related to net primary productivity patterns of the major North American natural vegetation formations. Regions of intense cultivation were observed as anomalous areas in the integrated growing season measurements. These anomalies can be explained by contrasts between cultivation practices and natural vegetation phenology. Major new information on seasonality, annual extent and interannual variability of vegetation photosynthetic activity at continental and global scales can be derived from these satellite observations.
Introduction
Vegetation is a key element in the environment and ecology of the earth. Full understanding of global climate, biospheric productivity, human impact on environment and related biophysical phenomena requires detailed knowledge of the geographic and seasonal patterns of terrestrial vegetation (Shukla & Mintz, 1982; Hansen el al., 1981; Woodwell el al., 1978; Sharpe, 1975; Thomas, 1959). Currently, information concerning the distribution and seasonality of global vegetation are derived from a wide range of information sources, such as maps, atlases, and field reports, which are of varying reliability and usually compiled from observations taken at different times. Recent efforts to estimate the global extent of selected vegetation types have produced results which differ by more than a factor of two for comparable vegetation types (Ajtay, Ketner & Duigneand, 1979; Olson & Watts, 1982; Matthews, 1983). Much less information is available on the global patterns of vegetation seasonality (Junge & Czeplak, 1968; Lieth, 1974; Fung et al., 1983). There is a need for consistent, timely and reliable information sources to facilitate analysis of global vegetation patterns. Remotely sensed spectral measurements of reflected solar radiation may contribute such an information source (Shay, 1969; NASA/HQ, 1983). This report discusses preliminary results of a study of North American vegetation patterns observed with visible and near-infrared measurements from the NOAA-7 Advanced Very High Resolution Radiometer (AVHRR).
Background
Spectral vegetation indices
Research carried out since the 1940's has shown that photosynthetically active 'green' vegetation displays a unique spectral reflectance pattern in the visible and near-infrared spectral regions when compared to other earth surface materials (Krinov, 1947; Colwell, 1956; Gates et al., 1965; Knipling, 1970; Colwell, 1974). The spectral reflectance of rocks, soils, dormant vegetation and woody plant materials, although variable in overall brightness; generally increases monotonically with increasing wavelength in the 0.4-1.0 um wavelength region. Photosynthetically active plant components, primarily leaves, produce a stepped reflectance pattern with low reflectance in the visible and high reflectance in the near infrared. This green vegetation spectral reflectance pattern results from strong absorption of visible light by chlorophylls and related pigments and scattering, because of leaf structural properties, but minimal absorption of light in the near infrared. Researchers have proposed a number of spectral vegetation indices premised on the contrasts in spectral reflectance between green vegetation and background materials (Rouse et al. 1974; Kauth & Thomas, 1976; Richardson & Wiegand, 1977; Tucker, 1979; Jackson, 1983). All of the indices are computed, at least in part, by calculating a difference or ratio of visible to near infrared measurements. This calculation minimizes the effects of variable background brightness while emphasizing variations in the measurements that occur because of varying green vegetation density. The normalized difference vegetation index (NDVI) is representative of the various spectral vegetation indices (Rouse el al., 1974).1t is computed;
NDVI = (NIR-VIS)/(NIR + VIS)
where NIR = near infrared spectral measurement
VIS = visible spectral measurement
In theory NDVI measurements range between -1.0 and +1.0. However, in practice the measurements generally range between -0.1 and +0.7. Clouds, water, snow and ice give negative NDVI values. Bare soils and other background materials produce NDVI values between -0.1 and + 0.1. Larger NDVI values occur as the amount of green vegetation in the observed area increases. The relative merits of the various spectral vegetation indices is still subject to discussion (Tucker, 1979; Jackson, 1983; Hatfield, 1983). However, analytical evaluation has shown that the various indices are functional equivalents (Perry Jr. & Lautenschlager, 1984).
The physical significance of spectral vegetation indices is under intensive investigation. Field studies. predominantly for agricultural crops and grasslands, have shown that the spectral vegetation indices correlate in a nonlinear manner with the green leaf area index and green biomass in the canopy (Dethier, 1974; Deering et al., 1974; Blair & Baumgardner, 1977; Holben et al., 1980; Bauer et al., 1981; Curran, 1983). These relations however are species-specific and saturate at high LAI and green leaf biomass levels. Theoretical studies of vegetation canopy radiance characteristics show that at a minimum the leaf area index (LAI), leaf angle distribution and leaf optical properties of the canopy must be known to predict observed canopy reflectance patterns (Idso & deWit, 1970; Suits, 1973; Bunnik, 1978). Parallel research on photosynthesis has shown that calculation of intercepted photosynthetically active radiation (IPAR) requires equivalent specification of canopy attributes (Weinman & Guetter, 1972; Lemeur & Blad, 1974; Monteith, 1977; Hesketh & Jones, 1980). Recent field studies of several different agricultural crops have shown a strong linear relation between IPAR and the spectral vegetation indices (Kumar & Monteith, 1981; Daughtry et al., 1983; Asrar et al., 1984). This IPAR-spectral vegetation index relation provides a physical link between the indices and plant productivity which is supported by studies of crops and grasslands which show a direct relation between the integrated value of spectral vegetation index measurements, acquired several times through the growing season, and the seasonal accumulation of biomass (Tucker et al., 1981; Daughtry et al., 1983; Hatfield, 1983; Steven el al., 1983; Tucker et al., 1985c). This research suggests that periodic visible and near-infrared observations of the earth may be used to characterize the current status, seasonal dynamics and integrated seasonal magnitude of vegetation photosynthetic activity.
Advanced Very High Resolution Radiometer (AVHRR)
Satellite-based remotely sensed observations have provided major impetus to vegetation studies based on spectral measurements. Satellites conveniently provide consistent, repeated coverage of all land areas. Much attention has been given to Landsat multispectral scanner (MSS) observations and selected studies have successfully used these measurements to analyze the regional extent, seasonality and productivity of natural and cultivated vegetation (Dethier, 1974; Deering et al., 1975; NASA/JSC, 1979; Thompson & Wehmanen, 1979). However Landsat data are not well suited for global scale studies. The MSS 80 meter ground resolution produces a large data volume over 6 000 MSS scenes are required for one observation of the earth's total land area and in cloudy regions of the globe, the 18 day repeat cycle produces too few cloud-free observations to record vegetation seasonal dynamics. The AVHRR sensor on the TIROS-N series of meteorological satellites, beginning with NOAA-6 launched in 1979, provides an alternate source of satellite visible and near-infrared measurements. With a 1.1 km nadir ground resolution and a daily repeat cycle, the AVHRR observations are well suited for large area studies. Several researchers have begun to use this new source of spectral measurements in vegetation studies and the results are encouraging (Townshend & Tucker, 1981; Gatlin et al., 1981; Gray & McCrary, 1981; Greegor & Norwine, 1981; Ormsby, 1982; Justice, 1983; Tucker et al., 1985a - c).
The AVHRK is a multispectral imaging sensor that was designed to permit detection and discrimination between clouds, land, water, snow and ice (Schneider & McGinnis, 1977). Spectral measurements are acquired in the visible, near infrared, middle infrared and thermal infrared regions, with four channels on the earlier AVHRR instruments on TIROS-N and NOAA-6 satellites and five channels on NOAA-7 and NOAA-8 (Table 1). AVHRR observations are available from the U.S. National Oceanic and Atmospheric Administration (NOAA) in several forms including High Resolution Picture Transmission (HRPT), Local Area Coverage (LAC), Global Area Coverage (GAC) and Global Vegetation Index (GVI) (Kidwell, 1979). The HRPT data are direct transmission data to ground receiving stations. The LAC data are the original 1.1 km observations, which are only recorded and placed in archives on special request. The GAC data, which are regularly recorded and stored, are partially resampled LAC observations. GAC data are produced by calculating the average value of the first four pixels in a 3 X 5 array of LAC observations. The average value is then used to represent the 3 x 5 pixel area. The Global Vegetation Index data are weekly summaries of channel 1 and channel 2 GAC data which have been geographically registered on a polar stereographic map projection and processed to minimize cloud cover and atmospheric haze (Tarpley et al., 1983). One grid cell of the GVI data represents ca. a 15 X 15 km ground area at the equator and increases in size with latitude to ca. a 30 x 30 km ground area at the poles. For each day of observations one GAC data value is randomly selected from the 10 to 60 GAC pixels present within each of the GVI grid cells to represent that GVI cell. The normalized difference vegetation index is computed for each of the selected seven days measurements. Measurements from the day with the largest NDVI value are selected to represent the composite weekly value. The NDVI value for that day is recorded, along with the associated channel 1 and channel 2 data, as the GVI data. Selection of the maximum NDVI value minimizes the effects of clouds and haze and tends to emphasize near nadir rather than off nadir observations (Holben & Frazer, 1984).
Data and methods
Two aspects of the North American AVHRR NDVI measurements are examined in this study; 1) seasonal variability and 2) the area under the curve (integral) defined by the temporal variation of the measurements over the growing season. Given current knowledge of spectral vegetation index measurements, the seasonal variations should be related to the phenology of vegetation green leaf display and hence absorbed photosynthetically active radiation and the integral of measurements over the growing season is expected to be related to annual net primary productivity (Monteith, 1977). However these relations have not previously been examined at a continental scale, across the diversity of vegetation types encountered in North America. The degree to which spectral vegetation index measurements provide a general indicator of vegetation activity, independent of the species or species composition observed, is unknown.
AVHRR Global Vegetation Index data were selected for this study for convenience since considerable data processing is already accomplished and observations of the entire North American continent are contained in a 1024 x 1024 geographically registered grid. Observations from NOAA-7 are used because the nominal daylight overpass time is 1430 hours compared to 0730 hours for NOAA-6 and NOAA-8. The afternoon solar illumination conditions provide more uniform and intense radiance measurements than the low zenith angle solar radiation observed in the early morning overpasses. A disadvantage of the afternoon observations is that cloud cover is usually greater at this time because of daytime atmospheric convection. The analysis presented in this report examines observations from April to November in 1982. Preliminary inspection of the winter (November to March) North American observations has revealed that low irradiance measurements near the polar terminator produce anomalous NDVI values. Restricting the current analysis to April through November avoids much of this problem while capturing the majority of the North American growing season north of 35deg. N latitude. South of 35deg. N and along the central west coast of the continent the full growing season is not observed in this analysis. All processing of the data was carried out on the HP-1000 minicomputer of the Sensor Evaluation Branch at the Goddard Space Flight Center.
Three-week composites
Seasonal variations in the spectral vegetation measurements were extracted from the data by forming three-week composites of the observations. Experience gained in analysis of NOAA-7 AVH RR data for the African continent has shown that at least three weeks of AVHRR observations must be composited to effectively remove cloud observations in cloudy regions of the continent (Tucker et al., 1985b). The compositing follows procedures previously described for the NOAA Global Vegetation Index data; that is, for each grid cell in the data, the week with the highest NDVI value is selected to represent the three-week interval. The resultant composite map is taken to represent the state of spectral vegetation index measurements across the continent during that three week interval.
Thirty GVI weekly composites, extending from 12 April 1982 to 7 November 1982, were processed to form ten three-week composites. Examples of the three-week composites for April, June, August and October are provided in Figure 1. The NDVI value at each grid cell location is color-coded, dependent on its magnitude, as shown by the color bar in Figure 1. The same color code is used for all the maps to permit interseasonal comparisons. A latitude-longitude coordinate grid, national boundaries and state boundaries in the United States are superimposed on the image to aid interpretation.
Integrated NDVI measurements
The area under the temporal curve was computed by trapezoidal integration of the three-week interval composite values at each grid cell location on the map. This resultant measurements are displayed in Figure 2. The same color code used in Figure 1 is used here with the exception that each value is multiplied by ten. These measurements are referred to as integrated NVDI ([[integral]]NDVI) values.
Results and discussion
Seasonal NDVI patterns
The three-week composites (Fig. 1) show that in general the NDVI measurements increase to a maximum in late August and thereafter decline. This seasonality is in agreement with the phenology of North American vegetation (Lieth, 1974). High NDVI values move northward across the continent in the spring and summer and shift to the south in the fall. This green-wave and brown-wave phenomenon has previously been studied for selected regions of the continent with Landsat observations where it has been related to the seasonal pulse of photosynthetic activity in the vegetation present (Dethier, 1974; Deering et al., 1975; Blair & Baumgardner, 1977).
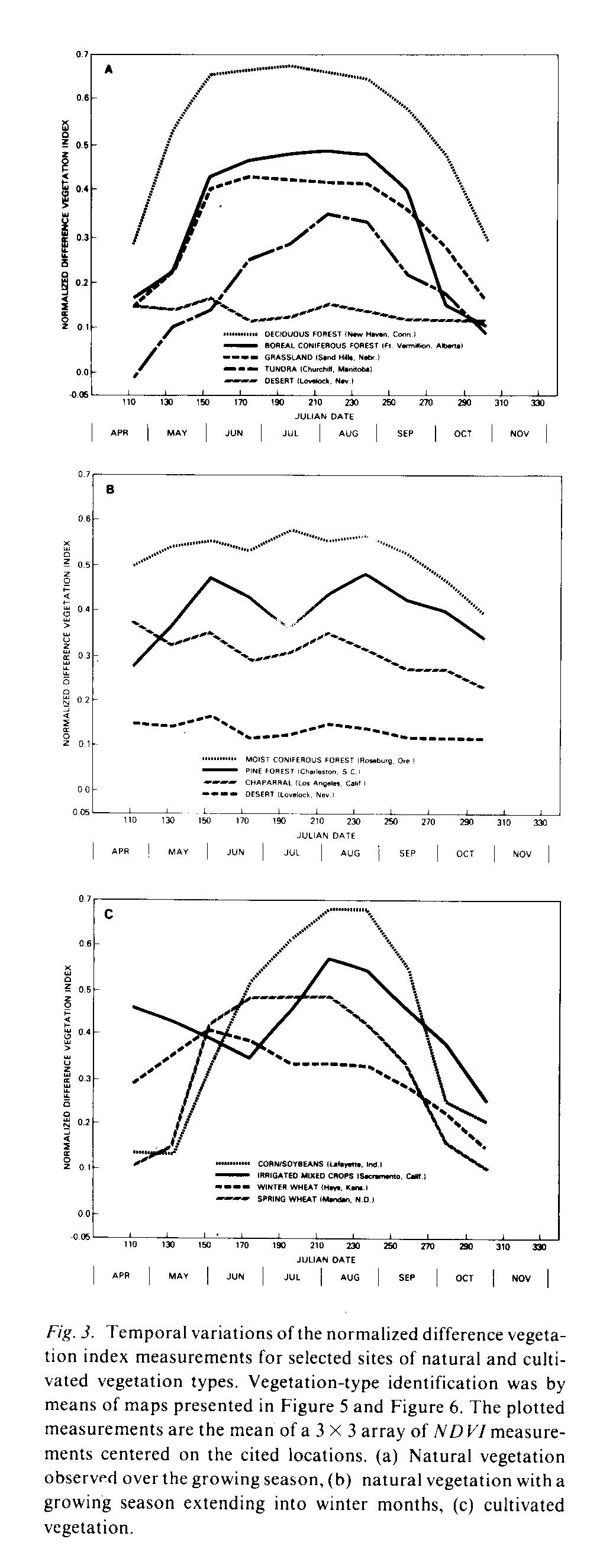
The seasonal variations of NDVI measurements for regions of selected natural and cultivated vegetation types are presented in Figure 3. These plots are the average of a 3 X 3 array of GVI three-week composite measurements, centered on the locations given. The observations are primarily from land dominated by natural vegetation although the effects of human activities in these locations cannot be entirely overlooked (e.g., irrigated agriculture). The plots in Figure 3a are for locations where the seven months of AVHRR observations cover the majority of the growing season. The Lovelock, Nevada observations of desert conditions are provided for comparison. As previously noted the measurements increase in the spring and decline in the fall. The magnitude of the summer measurements in general decreases with increasing latitude. Also, the duration of high summer values decreases to the north. However the grasslands observations from Nebraska peak at lower summer values than the boreal coniferous forest but with a longer duration of elevated values. These trends in the NDVI measurements correspond with the observed phenological patterns of these vegetation formations (Lieth, 1974).
The plots in Figure 3b are for regions of the continent where the full growing season is not observed in this seven month period. The NDVI plots for both Oregon and California show high NDVI measurements in April suggesting a continuation of vegetation activity from the winter months. The California observations steadily decline throughout the observation period which agrees with the increased vegetation dormancy observed in this region through the arid summer. The Oregon observations remain at high values until September and thereafter decline, reflecting the more humid character of this region as well as the physiology of these coniferous forests (Waring & Franklin, 1979). The South Carolina observations, from a region of pine forests, show a spring increase in the NDVI measurements and a small decline in the fall suggesting continued photosynthetic activity in the winter. No simple explanation of the mid-summer decline in NDVI values in South Carolina can be given. Several possibilities, including drought conditions in this region during 1982, cloud cover contamination, and atmospheric water vapor effects, may explain or contribute to an explanation of this pattern.
Representative temporal plots of NDVI measurements from prime agricultural regions are presented in Figure 3c. Both the corn-soybeans and spring wheat regions green up later and senesce (and/or are harvested) earlier than local natural vegetation. The winter wheat observations show high values in April given. The observations are primarily from land dominated by natural vegetation although the effects of human activities in these locations cannot be entirely overlooked (e.g., irrigated agriculture). The plots in Figure 3a are for locations where the seven months of AVHRR observations cover the majority of the growing season. The Lovelock, Nevada observations of desert conditions are provided for comparison. As previously noted the measurements increase in the spring and decline in the fall. The magnitude of the summer measurements in general decreases with increasing latitude. Also, the duration of high summer values decreases to the north. However the grasslands observations from Nebraska peak at lower summer values than the boreal coniferous forest but with a longer duration of elevated values. These trends in the NDVI measurements correspond with the observed phenological patterns of these vegetation formations (Lieth, 1974).
The plots in Figure 3b are for regions of the continent where the full growing season is not observed in this seven month period. The NDVI plots for both Oregon and California show high NDVI measurements in April suggesting a continuation of vegetation activity from the winter months. The California observations steadily decline throughout the observation period which agrees with the increased vegetation dormancy observed in this region through the arid summer. The Oregon observations remain at high values until September and thereafter decline, reflecting the more humid character of this region as well as the physiology of these coniferous forests (Waring & Franklin, 1979). The South Carolina observations, from a region of pine forests, show a spring increase in the NDVI measurements and a small decline in the fall suggesting continued photosynthetic activity in the winter. No simple explanation of the mid-summer decline in NDVI values in South Carolina can be given. Several possibilities, including drought conditions in this region during 1982, cloud cover contamination, and atmospheric water vapor effects, may explain or contribute to an explanation of this pattern.
Representative temporal plots of NDVI measurements from prime agricultural regions are presented in Figure 3c. Both the corn-soybeans and spring wheat regions green up later and senesce (and/or are harvested) earlier than local natural vegetation. The winter wheat observations show high values in April with a peak in June and declining values thereafter. The California Central Valley observations are high in April and decline until early July when they again increase to a maximum in late August. These temporal patterns are related to the calendar of cultivation and growth of crops which differs from natural vegetation phenology. This differential timing between cultivated and natural vegetation growth can also be observed in the three-week composites where the prime cropland areas display distinctive patterns in the images. For example the winter wheat region, extending from north central Texas through central Oklahoma and Kansas, stands out as an island of high NDVI values in the April image whereas the cornbelt shows up as an area of low values in this same image.
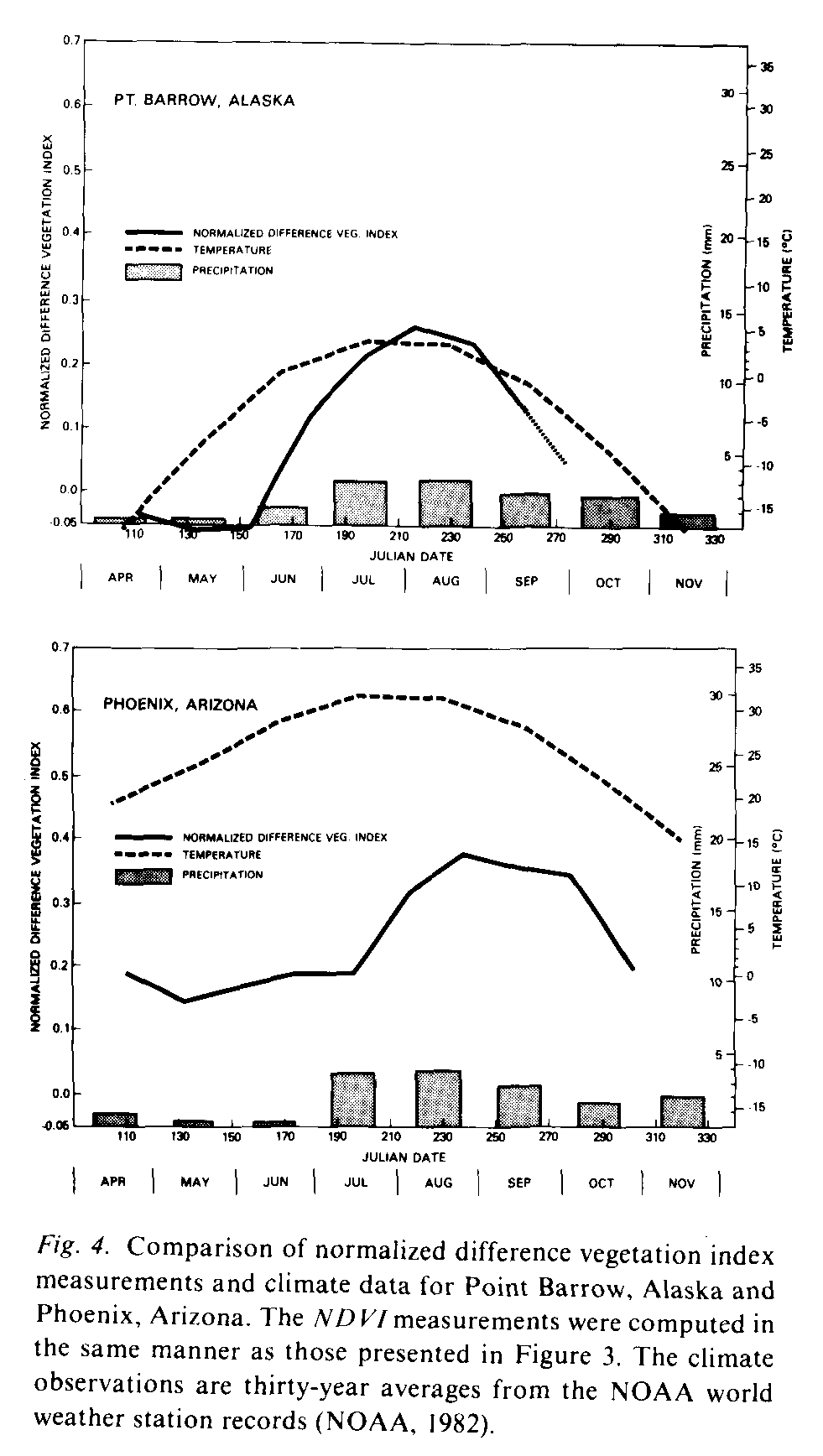
The NDVI measurements and climatic conditions should be related if the NDVI measurements are a sensitive indication of vegetation activity (Holdridge, 1947; Mather & Yashioka, 1968; Odum, 1971; Box, 1981). The NDVI measurements, precipitation and temperature records for Point Barrow, Alaska and Phoenix, Arizona for the seven month period are plotted in Figure 4. At Point Barrow the NDVI measurements only increase above 0.0 after mid-June, when the temperatures rise above 0deg. C, a condition essential for vegetation growth. In the Phoenix, Arizona region temperatures are always above freezing point but moisture limits vegetation growth. NDVI measurements for Phoenix follow moisture seasonality, showing lower values in the dry season from April to July and an increase in late summer when rainfall increases. This interaction between the NDVI measurements and climatic conditions is suggestive of the value of spectral vegetation index measurements in regional-to-global-scale vegetation phenology research.
Integration over the growing season
The map of integrated normalized difference vegetation index ([[integral]]NDVI) measurements (Fig. 2) displays a pattern of marked familiarity. The highest values occur in the southeastern and central west coast regions of the continent. The integrated measurements generally decrease to the north and west across North America but display more heterogeneity in the western one third of the continent. The north-south gradient of [[integral]]NDVI measurements corresponds to the major temperature ecocline of North America, whereas the east-west gradient corresponds to the major precipitation ecocline of the continent (Whittaker, 1970). The geographic complexity of the measurements in western North America is related to the mountainous terrain of the region. Note that elevation affects the measurements differentially dependent on latitude. North of 50deg. N latitude the [[integral]]NDVI measurements decrease with increasing elevation. South of 40deg.N latitude the measurements increase with increasing elevation. This geography of [[integral]]NDVI measurements compares well with the known patterns of North American natural vegetation activity (Shelford, 1963; Kuchler 1964; Whittaker, 1970; Odum, 1971; Rodin et al. 1975; Lieth, 1978).
The pattern of [[integral]]NDVI values in certain areas of the continent, particularly in the United States, appear 'anomalous' considering the natural vegetation and terrain of these locations. A region in the midwestern United States, extending from central Ohio to Iowa and including much of Indiana and Illinois, as well as portions of Wisconsin and Michigan, produce [[integral]]NDVI values that are lower than might at first be expected. A similar area of relatively low [[integral]]NDVI values extends north-south in the Mississippi River Valley from extreme southeastern Missouri through Arkansas and Tennessee to northwestern Mississippi. Conversely selected regions in the western United States, including the Columbia and Snake river basins in the Northwest, the Central and Imperial valleys in California and several locations in the Great Plains, exhibit higher than expected [[integral]]NDVI values. These locations correspond to the prime croplands in the United States as given by the World Atlas of Agriculture (1968). This map helped to explain the 'anomalous' areas observed in the integrated NDVI map (Fig. 2). The relatively low [[integral]]NDVI values in the U.S. Midwest occur because this region produces low NDVI values both early and late in the growing season, prior to planting and following harvest. Most agriculture in the western portion of the U.S. depends upon irrigation to sustain plant growth which produces greater plant cover and thus higher [[integral]]NDVI values than observed from natural vegetation of this semiarid region. These patterns correspond to the observed impact of agriculture on regional net primary productivity (Whittaker & Likens, 1975).
[[integral]]NDVI measurements and net primary productivity
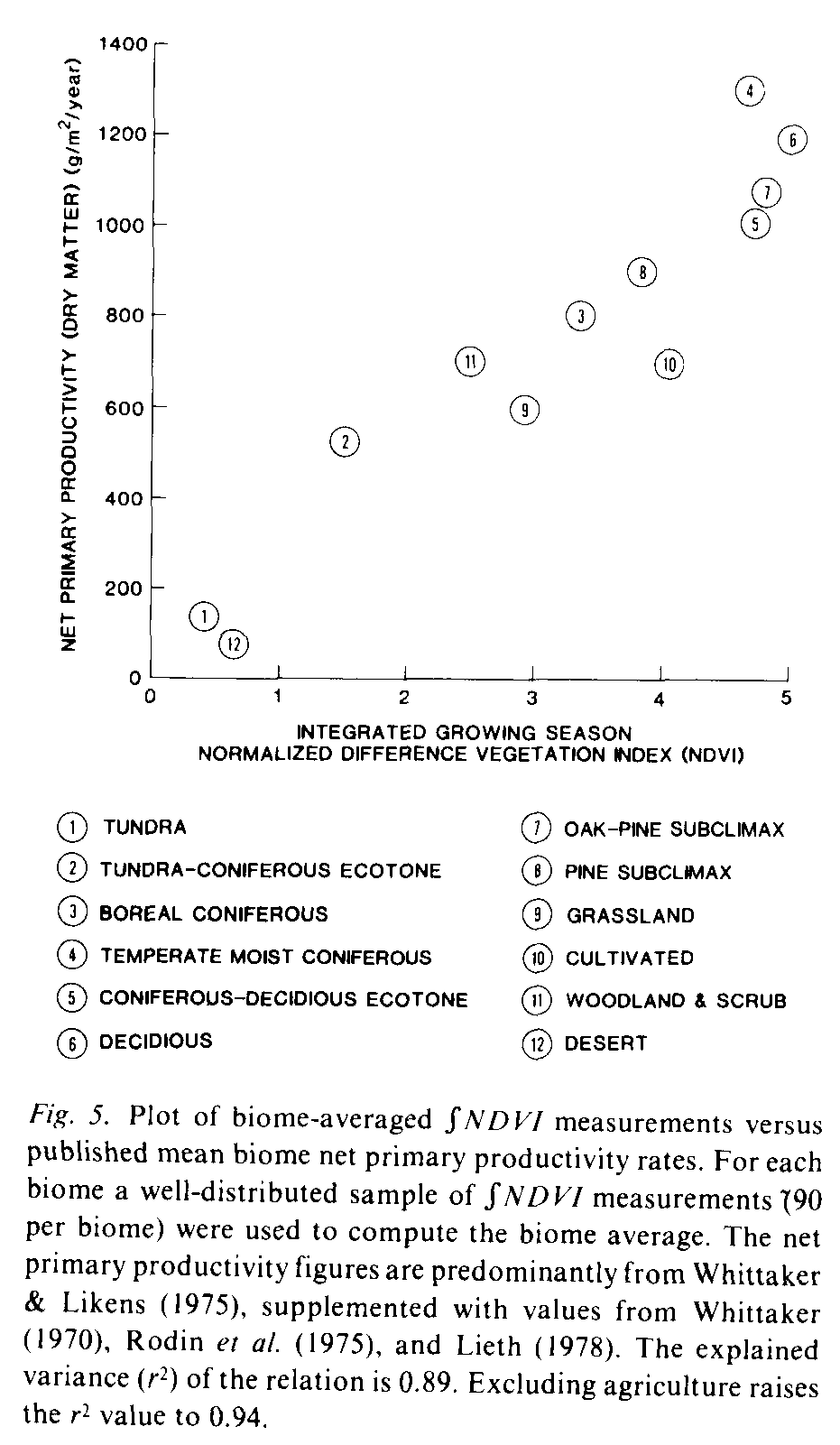
The relation between [[integral]]NDVI measurements and net primary productivity (NPP) was examined by computing the mean [[integral]]NDVI value for each of the major vegetation biomes of the continent and comparing these figures to net primary productivity figures reported in the literature. The North American biome map from Odum (1971) was used as a means to identify the natural vegetation formations below. This map was used to determine the natural vegetation types on the continent for the [[integral]]NDVI-net primary productivity analysis. The detailed western vegetation subregions were grouped to form the woodland and scrub category (sage subclimax, sagebrush, coastal chaparrel and pinon juniper) and the desert category (desert scrub and creosote-bush desert). [[integral]]NDVI samples were selected from sites within each formation based on the location of world weather observing stations used by World Meteorological Organization for global studies (NOAA, 1982). These sites were selected since they are uniformly distributed, approximately every 5deg. of latitude and longitude. across the continent. There are an average ten stations within each major biome. For each station the mean of a 3 X 3 pixel array of [[integral]]NDVI measurements, centered on the station location, was computed. The mean of all sites within each biome was then computed. These biome-averaged NDVI measurements were compared to net primary productivity figures provided by Whittaker & Likens (1975) supplemented by NPP figures for subclimax pine and pine/oak forests from Whittaker (1970) and values for the ecotones from Lieth (1978) and Rodin et al. (1975). The results (Fig. 5) show a strong direct relation between the NDVI measurements and net primary productivity. Excluding agriculture the explained variance (r2) of the relation is 0.94. With agriculture included the r2 value drops to 0.89. Further investigation is required before these results are fully understood. However the correspondence between these two variables is sufficient to suggest that satellite-derived spectral vegetation index measurements will significantly improve global studies of vegetation productivity.
Conclusions
The results from this analysis of NOAA-7 AVHRR observations of the North American continent show that these high temporal frequency, large area spectral measurements of the Earth provide a major new source of information for investigations of terrestrial vegetation characteristics. Seasonal variations in the NDVI measurements agree with the known phenological patterns for natural and cultivated vegetation and, for natural vegetation, these temporal patterns vary in concert with climatic conditions which limit plant growth. Annually integrated patterns of NDVI measurements correspond to known continental patterns of net primary productivity. The effects of human cultivation are clearly noted and these patterns agree with the known effects of agriculture on regional vegetation activity. Similar results have been derived for other continents (Tucker et al., 1985c; Justice et al., 1985). Use of AVHRR observations in macroscale studies of vegetation should significantly improve knowledge of global vegetation dynamics. Availability of several years of these high temporal frequency, globally consistent measurements will permit studies of interregional, interannual and seasonal vegetation phenomena not previously possible.
Much remains to be accomplished before the full value of the observations is realized. For example, interannual analysis of the data is needed and the effects of atmospheric conditions and sensor configuration on these measurements requires further investigation. However, even at this early stage of investigation, the results suggest that spectral vegetation index measurements provide a consistent and generalized means to conduct global vegetation studies. The AVHRR observations represent a major advance in realization of the promise of remotely sensed spectral observations for vegetation research because they provide the global overview of terrestrial conditions needed in multistage analysis of land conditions. An ability to observe the global distribution and dynamics of vegetation activity opens numerous new avenues of research for geographers, ecologists, climatologists and other earth scientists.
* This research is in part supported through NASA Cooperative Agreement NCC 5-26 from the NASA/Goddard Space Flight Center.
References
Ajtay, G. L., Ketner, P. & Duigneand, P., 1979. Terrestrial primary production and phytomass, In: B. Bolin, E. T. Degans, S. Kempe & P. Ketner (eds.) The global carbon cycle, pp. 129-181, SCOPE Report 13, John Wiley, New York.
Asrar, G., Fuchs, M., Kanemasu, E. T. & Hatfield, J. L.., 1984. Estimating absorbed photosynthetic radiation and leaf area index from spectral reflectance in wheat. Agro. 1. 76: 300-306.
Bauer, M. E., Daughtry, C. S. T. & Vanderbilt, V. C., 1981. Spectral-agronomic relations of corn, soybeans, and wheat canopies. Tech. Report 091281. LARS-Purdue University, West Lafayette, Indiana.
Blair, B. O. & Baumgardner, M. F., 1977. Detection of green and brown wave in hardwood canopy covers using multi-date, multispectral data from Landsat-I. Agro. J. 69: 808-811.
Box, E. O., 1981 . Macroclimate and plant forms: An introduction to predictive modeling in phytogeography. Tasks in Vegetation Science, I (H. Lieth, series editor). Dr. W. Junk, The Hague.
Bunnik, N. J. J., 1978. The multispectral reflectance of shortwave radiation by agricultural crops in relation with their morphological and optical properties. H. Veenman & Zonen, Wageningen, The Netherlands, 172 pp.
Colwell, J. E., 1974. Vegetation canopy reflectance. Remote Sensing Environ. 3: 175-183.
Colwell, R. N., 1956. Determining the prevalence of certain cereal crop diseases by means of aerial photography. Hildgardia 26 (5): 223-286.
Committee for World Atlas of Agriculture, 1968. World atlas of agriculture, Instituto Geografico de Agostini, Novara, Italy.
Curran, P. J .,1983. Multispectral remote sensing for the estimation of green leaf area index. Phi T Roy A 309: 257-270.
Daughtry, C. S. T., Gallo, K. D. & Bauer, M. E., 1982. Spectral estimates of solar radiation intercepted by corn canopies. Agristars Tech. Report SR-PZ-04236, Purdue University, West Lafayette, IN.
Deering, D. W., Rouse, Jr. J. W., Haas, R. H. & Schell, J. S., 1975. Measuring forage production of grazing units from Landsat MSS data. In: 10th International Symposium on Remote Sensing of Environment, ERIM, Ann Arbor, Michigan, pp. 1169-1178.
Dethier, B. E., 1974. Phenology satellite experiment. Final report, contract NAS5-21781, Cornell University, Ithaca, NY.
Fung, I., Prentice, K., Matthews, E., Learner, J. & Russell, G.. 1983. Three-dimensional tracer model study of atmospheric CO2: Response to seasonal exchanges with the terrestrial biosphere. J. Geophys. Res. 88: 1281-1294.
Gates, D. M., Keegan, H. J., Schleter, J. C. & Weidner, V. R., 1965. Spectral properties of plants. AP Optics 4: 11-20.
Gatlin, J. A., Tucker, C. J. & Schneider, S. R., 1981. Use of NOAA-6 channels one and two for monitoring vegetation, In: IEEE Proc. Int. Geoscience and Remote Sensing Symp., Washington, DC, June 8-11.
Gray, T. L. & McCrary, D. G., 1981. The environmental index, a tool potentially useful for arid land management. Agristars Rep. No. EW-NI-04076, NASA Johnson Space Center. Houston, Texas.
Greegor, D. H . & Norwine, J ., 1981. A gradient model of vegetation and climate utilizing NOAA satellite imagery, Phase I: Texas Transect. Agristars report no. FC-JI-04176, NASA Johnson Space Center, Houston, Texas.
Hansen, J., Johnson, D., Lacis, A., Lebedeff, S., Lee, P., Rind, D. & Russell, G., 1981. Climate impact of increasing atmospheric carbon dioxide. SCI 213: 957 966.
Hatfield, J. L., 1983. Remote sensing estimates of potential and actual crop yield. Remote Sensing Environ. 13: 301-311.
Hesketh, J.D. & Jones, J. W. (eds.). 1980. Predicting photosynthesis for ecosystem models. CRC Press, Boca Raton, Florida, 2 vols., 605 pp.
Holben, B. N. & Fraser, R. S., 1984. Red and near-infrared sensor response to off-nadir viewing. Int. J. Remote Sensing 5 145-160.
Holben, B. N., Tucker, C. J. & Fan, C. J., 1980. Assessing soybean leaf arca and leaf biomass with spectral data. Photogr. E. R. 46: 651-656.
Holdridge, L. R., 1947. Determination of world plant formations from simple climatic data. SCI 105: 367-368.
Idso, S. B. & deWit, C. T., 1970. Light relations in plant canopies. Appl. Opt. 9: 177-184.
Jackson, R. D.,1983. Spectral indices in n-space. Remote Sensing Environ. 13: 409-421.
Junge, C. E. & Czeplak, G., 1968. Some aspects of seasonal variation of carbon dioxide and ozone. Tellus 20: 422-434.
Justice, C. O., 1983. Contribution of remote sensing to the inventory and monitoring of natural resources. In: Proceedings, Renewable Resource Inventories for Monitoring Changes and Trends, Corvallis, Oregon, August 15 19: 362-366.
Justice, C. O., Townshend, J. R. G., Holben, B. N. & Tucker, C. J., 1985. Phenology of global vegetation using meteorological satellite data. Intern. J. Remote Sensing (in press).
Kauth, R. J. & Thomas, G. S., 1976. The tasseled cap a graphic description of the spectral-temporal development of agricultural crops as seen by landsat. Proceedings Symp. Machine Processing of Remotely Sensed Data. West Lafayette, Indiana.
Kidwell, K. A., 1979. NOAA polar orbiter users guide. Dept. of Commerce, Washington. D.C.
Knipling, E. B., 1970. Physical and physiological basis for the reflectance of visible and near-infrared radiation by vegetation. Remote Sensing Environ 1: 115-119.
Krinov, E. L., 1947. Spectral reflectance of natural formations. Akad. Nauk, USSR, Laboratorica Aerometodov, Moscow (Transl. by NEC of Canada, 11439, G. Helkov).
Kuchler, A. W., 1964. Potential natural vegetation of the Conterminous United States. Amer. Geographical Soc. Spec. Pub. 36, New York.
Kumar, M. & Monteith, J. L., 1981. Remote sensing of crop growth, In: H. Smith (ed.), Plants and the daylight spectrum, pp. 134-144. Academic Press, London.
Lemeur, R . & Blad, B. L., 1974. A critical review of light models for estimating the shortwave radiation regime of plant canopies. Agri. Meteorol. 14: 255 286.
Lieth, H., (ed.), 1974. Phenology and seasonality modeling, ecological studies 8. Springer-Verlag, New York.
Lieth, H., (ed.), 1978. Pattern of primary productivity in the biosphere. Benchmark Papers in Ecology, Vol. 8. Dowden, Hutchinson & Ross, Stroudsberg, PA.
Matthews, E., 1983. Global vegetation and land use; new high-resolution data bases for climate studies. J. Climate and Appl. Meteor. 22: 474-487.
Mather, J. R. & G. A. Yoshioka, 1968. The role of climate in distribution of vegetation. Assn Am. Geog. Ann. 58(1): 29-41.
Monteith, J. L., 1977. Climate and the efficiency of crop production in Britain. Phil. Trans R. Soc. Lond. B 291: 277-294.
NASA/GSFC, 1982 Project plan for fundamental research in scene radiation and atmospheric effects characterization. NASA/Goddard Space Flight Center, Greenbelt, Maryland, 128 pp.
NASA/HQ, 1983. Land-related global habitability science issues. NASA Headquarters, Washington, DC, 112 pp.
NASA/JSC, 1979. The LACIE Symposium, Proceedings of technical sessions. Vols. I and II, NASA Johnson Space Center, Houston, Texas, 1125 pp.
National Oceanic and Atmospheric Administration, 1982. Monthly climatic data for the world, sponsored by World Meteorological Organization. Vol.35, National Oceanic and Atmospheric Administration, National Climatic Center. Asheville, North Carolina. 172 pp.
Norwine, J. & Greegor, D. H., 1983. Vegetation classification based on AVHRR satellite imagery. Remote Sensing Environ. 13: 69 87.
Odum, E. P., 1971. Fundamentals of ecology. 3rd ed., W. B. Saunders. Philadelphia, 525 pp.
Olson, J. & Watts, J. A., 1982. Major world ecosystem complexes (map scale 1:30 m), included in W. C. Clark, (ed.), Carbon dioxide review 1982, Oxford University Press.
Ormsby, J. P., 1982. Classification of simulated and actual NOAA-6 AVHRR data for hydrologic land-surface feature definition, Geoscience and Remote sensing, GE-20: 262-267.
Perry Jr., C. R. & Lanternschlager, L. F., 1984. Functional equivalence of spectral vegetation indices. Remote Sensing Environ. 14: 169-182.
Richardson, A. J. & Wiegand, C. L., 1977. Distinguishing vegetation from soil background information. Photogr. E. R.43: 1541-1552.
Rodin, L. E., Bazilevich, N. I. & Razov, N. N., 1975. Productivity of the world's main ecosystems. In: D. E. Reichle, J. F. Franklin & D. W. Goodall (eds.), Productivity of the world's main eeosystems. National Aeademy of Sciences, Washington, DC.
Rouse, J. W., Haas, R. H., Deering, D. W. & Sehell, J. A., 1974. Monitoring the vernal advancement and retrogradation (Green wave effect) of natural vegetation. Final Rep. RSC 1978-4, Remote Sensing Center, Texas A&M Univ., College Station.
Schneider, S. & McGinnis, Jr., D., 1977. Spectral differences between VHRR and VISSR data and their impact of environmental studies. In: Proceedings Amer. Soc. Photogrammetry, 43 Meeting 27 February to 5 March 1977. Washington, DC.
Sharpe, D. M ., 1975. Methods for assessing the primary productivity of regions, In: H. Lieth & R. H. Whittaker (eds.), primary productivity of the biosphere, pp. 147-166, Springer-Verlag, New York.
Shelford, V. E., 1963. The ecology of North America. University of Illinois Press, Chicago, 722 pp.
Shay, R., (ed.), 1969. Remote sensing with special reference to agriculture and forestry. National Academy of Sciences, Washington, DC, 527 pp.
Shukla, J. & Mintz, Y., 1982. Influence of land-surface evapotranspiration on the earth's climate. SCI 215: 1498-1501.
Steven, M. D., Biscoe, P. V. & Jaggard, K. W., 1983. Estimation of sugar beet productivity from reflection in red and near infrared spectral bands. Intern. J. Remote Sensing 4: 325 334.
Suits, G. H., 1972. The calculation of the directional reflectance of a vegetative canopy. Remote Sensing Environ.2: 117-125.
Tarpley, J. D., Schneider, S. R. & Morey, R. L., 1983. Global vegetation indices from NOAA-7 meteorological satellite. J. Climate and Ap. Meteor. 23: 491-493.
Thomas, W. L., Jr., (ed.), 1959. Man's role in changing the face of the earth. The University of Chicago Press, Chicago, 817 pp.
Thompson, D. R. & Wehmanen, O. A., 1979. Using Landsat digital data to detect moisture stress, Photogr. E. R. 45: 201-207.
Townshend, J. R. G. & Tucker, C. J., 1981. Utility of AVHRR of NOAA-6 and -7 for vegetation mapping. In: Matching remote sensing technologies and applications proceedings, pp. 97-107, Remote Sensing Soc., London.
Tucker, C. J., 1978. A comparison of satellite sensors for monitoring vegetation. Photogr. E. R. 44: 1369-1380.
Tucker, C. J., 1979. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sensing Environ. 8: 127-150.
Tucker, C. J., Holben, B. N., Elgin, Jr., J. H. & McMurtrey, J. E., 1981. Remote sensing of total dry-matter accumulation in winter wheat. Remote Sensing Environ. 11:171-189.
Tucker, C. J., Gatlin, J. A., Schneider, S. R. & Kuchinos, M. A., 1982. Measuring vegetation in the Nile delta with NOAA-6 and NOAA-7 AVHRR data, In: Proceedings Intern. Symp. Remote Sensing of Arid and Semiarid Lands, Environmental Research Instituto of Michigan, Cairo.
Tucker, C. J., Vanpraet, C., Boerwinkle, E. & Gaston, A., 1984. Satellite remote sensing of total dry matter accumulation in the Senegalese Sahel. Remote Sensing Environ.13:461-474.
Tucker, C. J., Hielkema, J. U. & Roffey, J., 1985a. Satellite remote sensing of ecological conditions for desert locust survey and forecasting. Intern. J. Remote Sensing 6:122--138.
Tucker, C. J., Townshend, J. R. G. & Goff, T. E., 1985b. Continental land cover classification using NOAA-7 AVHRR data. SCI 227: 369 - 375.
Tucker, C. J., Vanpraet, C., Sharman, M. I. & Van Ittersum, G., 1985c. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel: 1980-1984. Remote Sensing Environ, 14: 233-249.
Waring, R. H. & Franklin, J. F., 1979. Evergreen coniferous forests of the Pacific Northwest. SCI 204:1380-1386.
Weinman, J. A. & Guetter, P. J., 1972. Penetration of solar irradiance through the atmosphere and plant canopies. J. Appl. Meteorol. 11:136-140.
Whittaker, R. H., 1970. Communities and ecosystems. Macmillan, London, 162 pp.
Whittaker, R. H. & Likens, G. E., 1975. Primary production: The biosphere and man, In: H. Lieth & R. H. Whittaker (eds.), Primary productivity of the biosphere. Ecological Studies, 14, Springer Verlag, New York.
Woodwell, G. M., Whittaker, T. H., Reiners, W. A., Likens, G. E., Delwiche, C. C. & Botkin, D. B., 1978. The biota and the world carbon budget. SCI 199:141-146.
{kind=link}
{kind=link}
{kind=link}
{kind=link}